1. Introduction
Barley
(Hordeum vulgare L.) is one of the most important cereals in the world. Based on cultivated area and production quantity, among the small grains, it places right after wheat and before oats, triticale, and rye, both globally and locally. In the Republic of Serbia, barley takes second place with a production of 470.000 t [
1] and an average grain yield in 2022 of 4.8 t ha
−1 [
2]. Although it is used in various ways, mainly as feed or as raw material in the beer industry [
3], cultivar selection and breeding are based primarily on grain yield, which represents a complex trait influenced by various factors [
4].
The weather conditions change unpredictably from year to year, making it difficult to predict grain yield and its stability for plant breeders [
5] and increasing income fluctuations for farmers [
6]. Temperature, amount and distribution of precipitation, and applied production technology significantly affect the grain yield to be lower than the genetic potential of cultivated varieties [
7]. Although it is less susceptible to the influence of drought stress compared to other small grain cereals (such as rye and wheat), abiotic stress is a major limiting factor for barley production [
8].
Multi-environment yield trials are usually conducted to obtain information regarding superior genotypes for cultivation and include two main factors: genotype (G) and environment (E). The environment encompasses locations, soil characteristics, sites, years, and agronomic practices [
9], which influence the performance and adaptability of different genotypes under varaying conditions. In trials, changes in the rank of genotypes from one environment to another, a difference in scale between environments, or a combination of these two situations can often occur. This is due to genotype by environment interaction (GEI), which, in addition to environmental and genotype effects, influences factors in the phenotype of traits [
10]. GEI can be defined statistically as the difference between the phenotypic, experimental assessment and the value predictable from the theoretical model of observations, which takes into account the general mean as well as genotypic and environmental main effects [
11]. Breeders intend to create adaptable genotypes that are less sensitive to changes in environmental conditions, ensuring consistent yield levels and minimizing their contribution to the GEI [
12].
GEI is a complex problem caused by numerous climatic, edaphic, genetic, morphological, phenological, and physiological variables and often cannot be explained. Knowledge about the influence of environmental factors on the growth and development phases of certain crops could improve the selection of stable genotypes and reduce the possibility of yield decrease in various environments [
13], while environmental parameters had the most significant impact on particular months [
14]. Understanding the genotypic responses to environmental factors can help in GEI interpretation and exploitation [
15].
Some statistical models can use external information directly to study GEI. Opposite to empirical models, these analytical models understand the morphophysiological and agromorphological causes of genotype response [
16,
17]. Partial least squares regression (PLSR) is a bilinear analytical statistical model that combines external variables (both environmental and genotypic) to study and interpret GEI [
18]. PLSR analysis is a powerful tool due to its ability to separate interaction from main effects using environmental condition parameters during the specific growth stages but also because it gives us the answer about the biological causes of the interaction [
19]. The advantage of PLSR over other statistical approaches is that it can identify only relevant predictor variables, while linear models require pre-selection of potential predictor variables before analysis and are analytically limited. When the number of variables is greater than the observations and there is high collinearity among variables, it is pragmatic to use the PLSR method [
20].
Results of several statistical models indicate that the PLSR model is effective in detecting the environmental variables that explain a sizeable proportion of GEI variability [
21] and that this model is also powerful for analyzing the influence of climatic variables on the biological phenomena variation [
22]. PLSR is a novel and robust method of handling complex cereal phenology and climatic data. Improving grain yield and adaptation is achieved by synchronizing crop phenology with the environment and identifying crucial periods during the growing season [
23]. Grain yield is formed continually from sowing to maturity. However, some phases and variables associated with them are more significant for the determination of the final yield [
24]. By knowing the response genotypes to these factors, we can predict the timing of the phenological phase with this model. For example, using the PLSR model, the anthesis is highlighted as an important phase for the adaptation of barley, and the temperature and day length are important variables for determining the anthesis [
23].
There are sparse data on the direct influence of climatic factors on barley yield using PLSR. The application of the PLSR model in cereals is mainly used on wheat, so data on barley are very scarce. The temperature is crucial for all phenophases, from sowing to maturity [
25], while the maximum soil temperature at 5 cm in April, May, and June (during spike growth, anthesis, filling, and maturity stage) and sun hours per day in May are the factors responsible for the interaction of grain yield in wheat [
26]. Minimum and maximum temperatures during the spike primordia stage (during March), as well as precipitation during the rapid spike growth and anthesis stage (during April and May), were found to be significant factors influencing bread wheat yield [
27], while in dry conditions, relative humidity had a special role in interaction across all periods [
15]. In durum wheat, it was reported that the maximum and mean temperatures during the entire crop cycle were the most important environmental factors influencing GEI [
28]. The most sensitive period in the development of bread wheat is the rapid spike growth stage [
26,
27], as well as the spike primordia growth stage in wheat and triticale [
19]. More recently, PLSR has found application in genomic selection where whole-genome markers are used to predict and describe a phenotype [
29] so that crop phenology can be used for genetic dissection. Apart from small grains, various authors have used this model on other field crops such as corn [
30], sunflower [
31], and soybean [
32].
Considering the importance and consequences of interaction in barley breeding, this study aimed to: (i) estimate GEI for grain yield in winter barley using a linear mixed model; (ii) determine the most important environmental factors at different stages of development that influence the GEI of barley grain yield by applying the PLSR model; (iii) assess cultivar differences in sensitivity to environmental factors at different crop stages; (iv) determine the relationships of variables in environments favorable for barley growing and predict grain yield stability in new environments with the help of weather data; (v) single out suitable test locations for barley selection when the environment represents a combination of location and years and to give recommended genotypes in these environments; and (vi) identify genotypes intended for cultivation in wider geographical areas, as well as those intended for specific areas (wide and specific adaptation).
4. Discussion
For the analysis of yield trial data from multiple environment trials, it is prevailing to assume a linear mixed model, where genotypes are fixed, and environments and interactions are random [
44], which is congruent with our study. The mixed model should be routinely used for genotype evaluation in multi-environment trials since it is considered superior compared to the classical ANOVA model [
45]. Since a heterogeneous mixed model was chosen in our study, this further implies that the heterogeneity of REV generally exists and indicates that the REV varied across the environments.
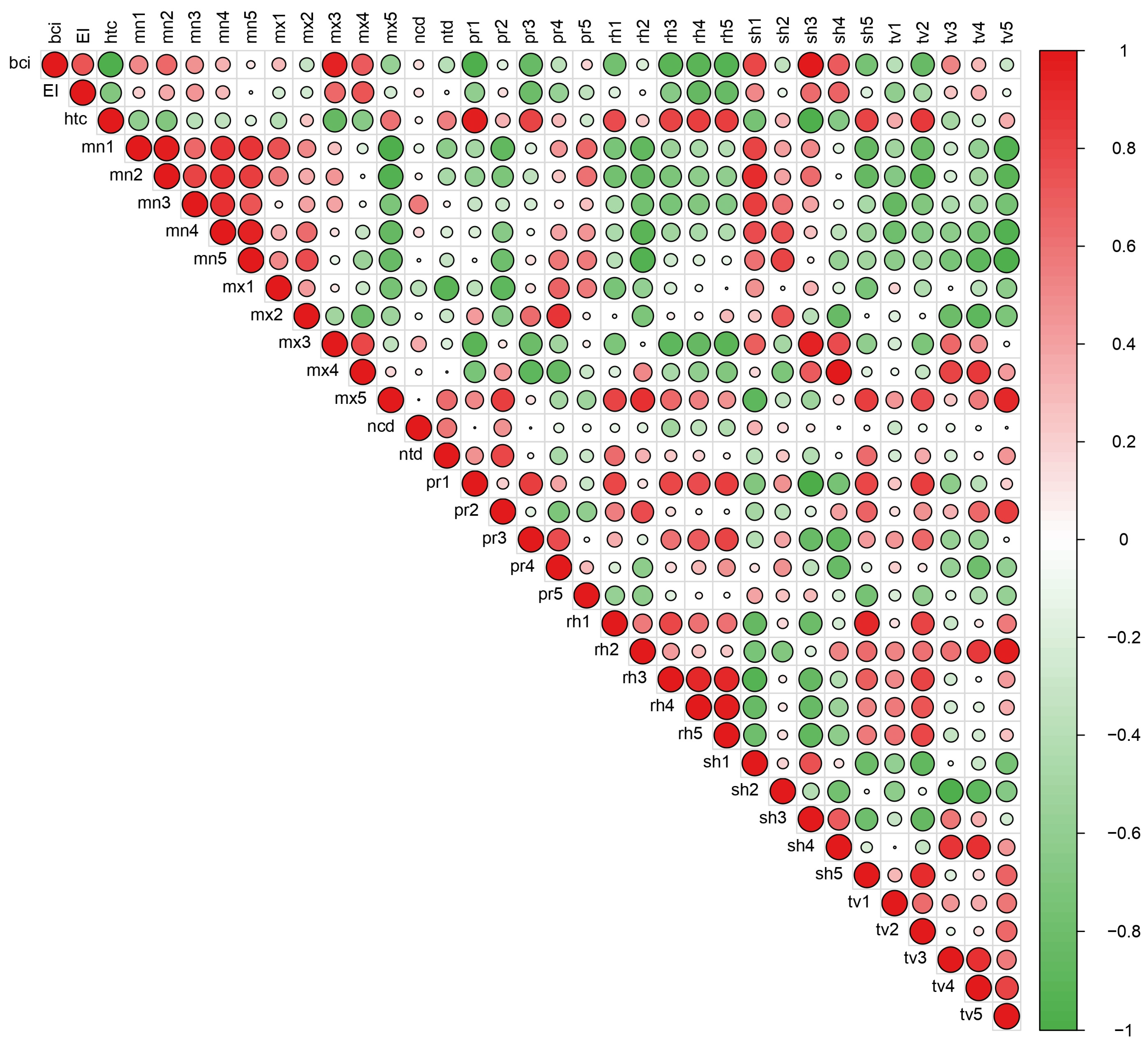
A sample of 20 barley genotypes, in terms of size, can be considered random and representative. The results presented in this paper reflect the complexity of interaction. The presence of GEI could be attributed to the differences among the genotypes and environmental conditions of the three sites across two years. Considering that the PLSR model explained a 43.7% sum of squares of interactions, for further analysis, it will be necessary to include the other variables that were not included in our research.
The PLSR method was shown to be effective in detecting environmental explanatory variables associated with factors that explained large portions of GEI. The highest loadings of variables indicated a high correlation with grain yield [
38]. Although the first dimension in our research explained the higher portion of interactions, the variables closely related to the second dimension cannot be ignored. According to Vargas et al. [
21], the second PLSR factor improves the prediction accuracy of the model, even if it is not significant.
In our study, the first and second dimensions were statistically significant, and thus, twenty variables related to both dimensions were singled out. A substantial contribution to the interaction of almost all examined environments was due to the large number of isolated variables, which concurs with research by Reynolds et al. [
19]. Kondić-Špika et al. [
27] singled out variables to explain the interaction based on the positive or negative relationship between the highest-yielding and the lowest-yielding environments. Considering that with the approach used by Kondić-Špika et al. [
27], variables rh1, tv1, tv2, sh5, and mx5 would not be included, we did not use it in our research. Our research on barley yield interactions showed different variables in all periods, i.e., phenophases of barley development, which indicates that grain yield is formed continuously from sowing to maturity [
24].
The variables emphasize the importance of precipitation and sun hours from November–February (pr1, sh1) when barley goes through germination, emergence, and the beginning of tillering (vegetative stage). Pržulj and Momčilović [
46] indicated that barley grain yield was positively correlated with precipitation and the sum of active temperatures in the vegetative period. Therefore, the high vegetative mass before flowering is a prerequisite for high yields. With regular winter precipitation, winter barley usually completes the vegetation before the first spring moisture deficit and successfully uses the moisture accumulated during the winter months. Water excess can negatively affect barley seed production, especially after germination [
47]. That concurs with our results and represents one of the reasons for the lower grain yield in the second growing season. Ploschuk et al. [
48] stated that depending on the waterlogging duration, phase development, soil type, and genotypes, reductions in barley yield range from 18 to 71%, while wheat is much more tolerant (between 14 and 29%). In our research, temperatures did not show significance. However, temperatures’ indirect effect through the sun hours was noticed. Using the PLSR model, Reynolds et al. [
19] singled out solar radiation as significant for the interaction in wheat in the vegetative phase. The negative impact on the yield of relative humidity from November–February (rh1) can be indirectly explained by the relationship with the amount of precipitation in this period.
Our study indicated that barley favors warmer night temperatures during March (mn2). In ecological conditions of Serbia, it is a period when the spike primordia stage takes place (tillering and beginning of stem elongation), and potential grain and spike numbers are determined at temperatures between 6 and 20 °C [
49]. Maximum temperatures (mx2) also showed significance, which is the reason for the positive effect on the yield of slight temperature variations (tv2) in this period. This concurs with the findings of Kondić-Špika et al. [
27] and Reynolds et al. [
19], who obtained similar results on wheat by applying the PLSR model, highlighting the importance of temperature variables. McMaster et al. [
50] emphasize the importance of the temperature in the double ridge phase on the apical meristem that takes place during tillering, which is the basis for potential spike fertility and grain yield.
The variables sh3 and mx3 had a significant influence on interaction and a positive effect on yield. During April, barley grows intensively in height and forms biomass, which is highly important for grain yield [
51]. Kondić-Špika et al. [
27] and Abbate [
52] suggested that bread wheat yield is the most sensitive to environmental conditions during the rapid spike growth stage (April in Serbian conditions). Calderini et al. [
53] indicate that solar energy at this stage influences kernel weight potential and that grain number is increased in conditions of lower average temperature and a higher number of sun hours. This partly concurs with our findings since more sun hours and maximum temperatures positively affected the yield and produced higher productivity of the first growing season compared to the second one.
The largest number of variables (mx4, tv4, sh4, pr4, rh4) showed their importance during May, when the anthesis and filling stage took place in the climatic conditions of Serbia. Porker et al. [
23] pointed out the importance of the period of anthesis on the yield and adaptation in barley, which is also shown by our research. This is the most active period for photosynthesis in barley, the basic physiological process of plant growth that provides the necessary energy, and when most of the organic matter of yield is produced. Favorable temperatures and enough light enable as many flowers as possible to be fertilized and as much matter as possible to be transported to the grain, which directly affects the yield [
54]. These conditions were also singled out by Dodig et al. [
26], who applied the PLSR model to study the interaction affecting wheat yield, which concurs with our investigation. Voltas et al. [
55] indicated the importance of temperatures during grain filling, while Kondić-Špika et al. [
27] reported a negative impact of low-temperature variations on the yield in May (tv4), which is highlighted in our research. The most negative influence of high temperatures occurs during anthesis and grain filling, which reduces the conversion of simple sugars into starch [
56]. High temperatures during the grain-filling period had an impact on yield by reducing grain number, grain weight, and grain quality [
57], while heat stress for five days in the middle of this period decreased the yield of barley by about 35% [
58]. Pržulj and Momčilović [
59] indicated that a combination of moderate temperatures and adequate rainfall levels during anthesis and grain filling enables the achievement of a high yield of barley. These authors define temperatures in the grain-filling period as moderately high when the maximum temperature is between 15 °C and 25–30 °C and very high, i.e., heat stress, when the maximum temperatures are between 35 °C and 40 °C. Based on that, during the experiments, the maximum temperatures were moderately high, but the second growing season had a period of ten days with much lower values (<10 °C). In our study, the reduction of the assimilation process and reduction of yields during the second growing season were also influenced by lower values of maximum temperatures (mx4), temperature variations (tv4), and sun hours (sh4). Arenas-Corraliza et al. [
60] indicated that barley shows better acclimation ability to shade compared to wheat. However, reduced light conditions cause lower photosynthetic activity and grain yield in general, which is confirmed by this study. Low light intensity decreases stem strength by restraining carbohydrate assimilation (especially lignin biosynthesis) and reducing culm-wall thickness [
61]. The relationship with lodging resistance in wheat and barley described by Li et al. [
62] and Yu et al. [
63] was one of the reasons why lodging was recorded in the second growing season of our research.
Both a lack and excess of and precipitation in this stage can negatively affect barley grain yield. Compared to other cereals, barley is well adapted to drought [
64], but the lack of accessible water in the grain-filling period strikes photosynthesis. This condition results in reduced accumulation of biomass, more abortive flowers and grains, a decrease in the mass of individual grains [
65,
66], acceleration of the grain-filling period [
67], and a reduction of grain yield in two-row barleys by 28.8% [
68]. High values of precipitation cause waterlogging and lodging of the stem. Our findings indicate that high precipitation in May (pr4) affected the significant interaction and yield reduction during the second season in environments KG10 and ZP10. This concurs with de San Celedonio et al.’s [
47] findings, which reported that barley is the most sensitive to waterlogging around the flowering phase.
Lodging is the state of permanent displacement of the stems from their upright position and is one of the main factors constraining grain yield stability in barley depending on the lodging intensity and the development stage of its occurrence. Caierão et al. [
69] indicated that the greatest lodging-induced reductions in potential grain yield in barley and other cereals occur when crops are lodged flat at anthesis or early on in grain filling (25–75%), which was confirmed by our research. Precipitation at the beginning of the filling stage affected the lodging and thus reduced yields in environment KG10. Smaller yield losses also occur when lodging occurs at a later stage of development, especially in maturity [
70]. This is probably one of the reasons why the precipitation in June was not significant for the interaction, even though, in the second growing season, there was lodging (ZP10) caused by precipitation in this period.
The lower the humidity is, the easier it is for the plant to release water since as the relative humidity of the air surrounding the plant rises, the transpiration rate falls. This reduces stomatal conductance for water vapor and gases and moves nutrients and water, resulting in a photosynthesis reduction [
71]. Under these conditions, the other vital plant processes are severely restricted, and as a result, developing flower growth and new growth are damaged. Csajbók et al. [
72] indicated that with the development of winter barley, the positive correlation between assimilation rate, transpiration rate, and stomatal conductance increases and reaches a maximum at the end of April and throughout May, when flowering and grain filling occur. This concurs with our observations since the variables rh3 and rh4 have shown significance for the interaction, and their high values had an adverse impact on the fertilization of flowers, as well as on the assimilation and transport of matter in the grain during the second growing season.
Sun hours, minimum and maximum temperatures, and relative humidity during June (sh5, mn5, mx5, and rh5, respectively) showed importance for barley maturity. Yield components of barley are formed during the whole vegetation, but kernel weight is formed during the period between anthesis and physiological maturity [
73]. Majoul-Haddad et al. [
74] stated that high-temperature stress causes earlier harvest maturity, producing shriveled grain and decreasing the yield of small grains by up to 50%. By applying the PLSR model for wheat yield during maturity, Dodig et al. [
42] found the significance of maximum soil temperatures (related to maximum air temperature and radiation), which was confirmed by our research.
Jacott and Boden [
75] reported the importance of considering high minimum and maximum temperatures in assessing the response of wheat and barley to a warming climate, particularly during the later productive stages. In the conditions of Serbia, it rarely happens that minimum temperatures influence the yield or maturity of barley before June, while Garcia et al. [
76] indicated that the lower the latitude is, the higher the grain yield losses are due to night temperature increase. The effect of high night temperatures may be correlated with a decrease in the amount of photoassimilates available for plant growth caused by higher respiration at night and accelerating the process of maturity, which leads to reductions in the final grain weight [
77].
The indirect effect of humidity is that damp, stagnant conditions encourage mold and bacterial diseases, which can lead to reduced yields throughout the growing season, especially during the filling and maturity of barley (rh4, rh5).
Our investigation points out the influence of the ratio of precipitation and temperature (climate humidity), as well as the ratio of precipitation, temperature, and sun hours during the whole vegetation, which is indicated by a hydrothermal coefficient and bioclimatic index (htc and bci, respectively). This concurs with the results of Voltas et al. [
55], who used the PLSR model to explain interactions and barley yield by the relationship between precipitation and temperature during the entire vegetation period. Vargas et al. [
21] did not find that precipitation affects the interaction, but only temperatures and sun hours.
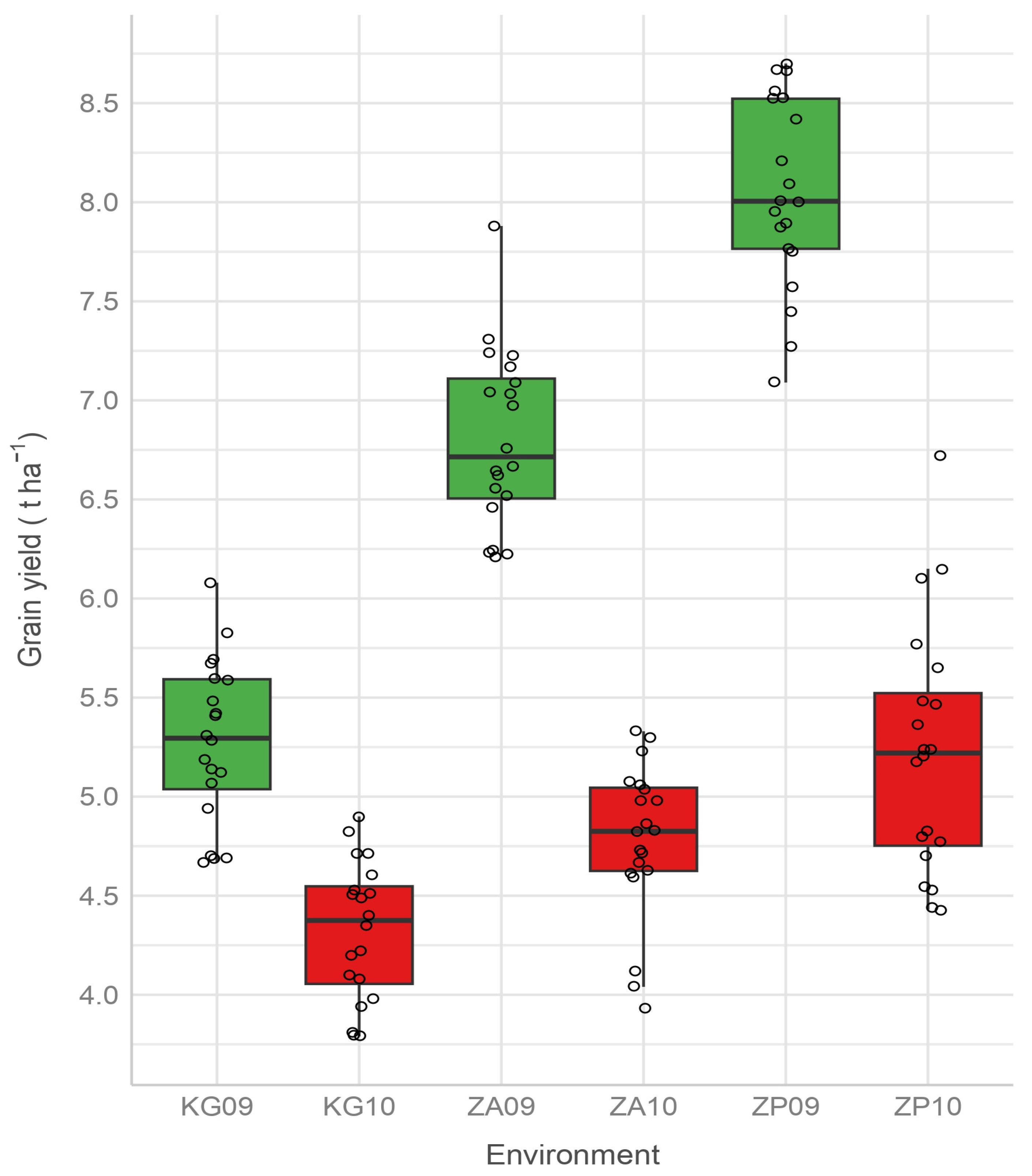
All the environments were low yielding and humid by the hydrothermal coefficient (htc > 2) in the second growing season. At the same time, they had lower values of the bioclimatic index. Reynolds et al. [
19] also pointed out that precipitation may be a characteristic of low-yielding environments. However, in the first growing season, ZA09 was a high-yielding and humid environment (htc > 2), unlike ZP09 (high yielding) and KG09 (low yielding), which were optimally humid (1 ≤ htc ≥ 2). The values of these two indices for high-yielding environments ZP09 and ZA09 were between 1.82 (optimally humid) and 2.41 (humid) for htc and between 2.44 and 1.71 for bci, so the optimal values for the growth and development of barley are in this range. The possible explanation why KG09 (1.94 and 2.21, respectively) belongs to the group of low-yielding environments is that edaphic conditions at KG09 are probably poorer than in other investigated environments.
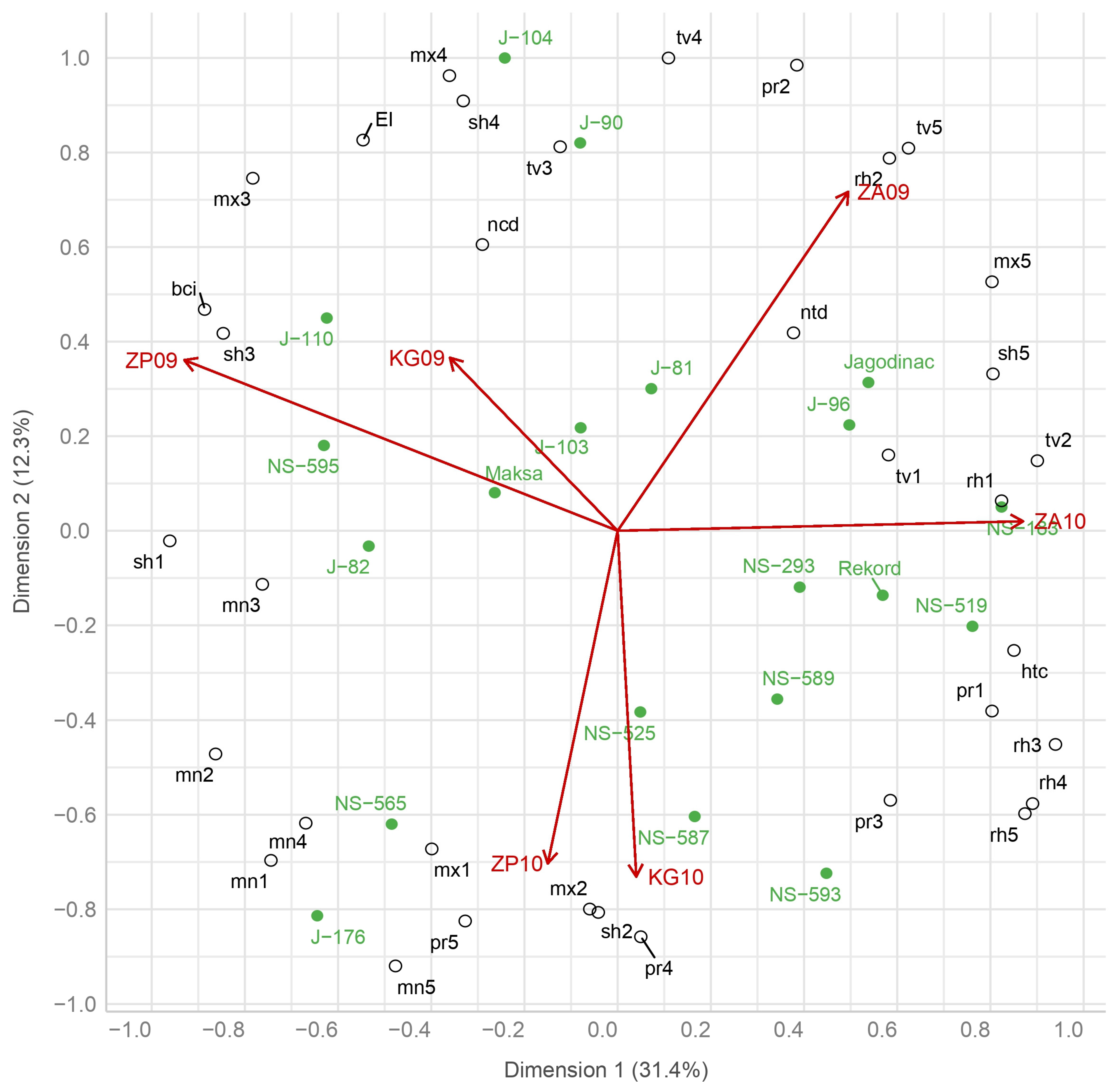
The results also showed that genotypes do not respond similarly to environmental variables at different stages of development, and therefore, we observed three clusters of genotypes. The first cluster consists of high-yielding genotypes NS-593, NS-565, and J-176 (two out of the three highest yielding) and one low-yielding genotype, NS-587. These genotypes represent younger genetic material created after 2000. They positively interacted with humid environments ZP10 and KG10 (recorded lodging of the stem). Based on the biplot display, it is suggested that these genotypes’ performances are associated with high precipitation during the spike growth stage onwards when barley is most sensitive to lodging, which can significantly reduce yields. This interaction can be partly explained by less sensitivity to lodging, which may be caused by lower stem height (all were low except J-176) or some other feature (wall thickness or more intense lignin production). Therefore, genotype J-176 may be a significant source of genes in further breeding due to its specific tolerance to lodging. These environments are also characterized by poor conditions for photosynthetic activity (negatively associated with temperatures and sun hours, as well as positively associated with relative humidity during April and May). However, the genotypes showed great stability regarding environmental effects on grain number and, especially, kernel weight determination in the spike growth and grain-filling stage. Csajbók et al. [
72] indicate a large diversity in barley in terms of assimilation rate in poorer conditions for photosynthesis, which may be the reason for the better adaptability of these genotypes. These conditions reduced the positive temperature effect for grain number determination during the spike primordia stage in March.
The second cluster represented a group more sensitive to these conditions and consisted of high-yielding genotypes NS-595 and J-82 and low-yielding genotypes J-104, J-90, and J-110. All these genotypes belong to younger genetic material released after 2000. They had a positive interaction with optimally humid environments (ZP09 and KG09). This suggested that these genotypes favor highly moderate temperature conditions and sun hours in which assimilates are created (during the spike growth and grain-filling stage in April and May, respectively), which indicates that high yield was driven principally by kernel weight. These genotypes mostly had high and medium heights of the stem (except for J-90 and J-110). However, it was assumed that the interaction was partly a consequence of more sensitivity to the lodging of genotypes (except for J-110) in conditions represented in humid environments (ZP10 and KG10).
The third cluster consisted of three of the lowest-yielding genotypes, Jagodinac, Rekord, and NS-293, as well as high-yielding NS-183, NS-519, and J-96, which had a positive interaction with both humid environments in Zaječar. Except for the breeding line J-96, all cultivars were released before 2000 and represent older genetic material. Performances of these genotypes were associated with high temperatures and sun hours during June, as well as lower minimum temperatures during the whole season. Presumably, these genotypes were well adapted to warmer conditions during maturity. The earliness did not show any significance since positive interactions were realized with early (Jagodinac, NS-183, and Rekord), later (J-96 and NS-519), and medium-late (NS-293) genotypes. Dodig et al. [
78] also confirms this claim by pointing out that two-row barley generally showed good adaptability to poorer weather conditions at the grain-filling and maturity stage, so that earliness is not as important as in six-row forms. These environments were characterized by low values of precipitation during these phases (pr4 and pr5), and therefore a lower probability of lodging, which explains the fact that the genotypes mostly had a taller stem (except NS-519 and Rekord). Therefore, better adaptability to these conditions of these genotypes we can explain by the high stem biomass. According to Savić et al. [
79], a taller stem is related to high stem biomass, which determines the potential for accumulation and remobilization of dry matter from stem to grain in stress conditions. Less sensitivity to cold conditions (especially night) in stages during winter and spring is considered as important regarding genes for frost resistance. These genotypes showed stability under relatively poor environmental conditions concerning grain and spike number determination in the spike primordia stage during March (negatively correlated with mn2 and mx2, and positively with tv2).
Most genotypes showed a positive interaction with low-yielding and humid environments. These environments have affected both the highest-yielding (NS-525, J-176, and NS-593) and the lowest-yielding (Jagodinac, NS-293, and Rekord) genotypes. The same results were reached by Dodig et al. [
78] and Hilmarsson et al. [
80], who indicated better adaptability of two-row barley in low-yielding environments compared to six-row barley, which is more favorable in high-yielding environments.
The study of interaction is the basis for genotype selection intended for cultivation in wider geographical areas and for those intended for specific areas [
81]. In our study, genotypes J-104, J-90, J-176, NS-565, and NS-593 had significant contributions to the interaction, which, according to Gauch [
82], indicated that these genotypes were under more substantial influence of the environment. On the other hand, genotypes that showed a smaller contribution to the interaction reacted better to changes in external factors and are intended for growing in wider geographical areas, thus indicating stability. Elakhdar et al. [
9] highlighted that in order to use the beneficial effects of the interaction and the selection of favorable barley genotypes to be more precise, the stability and average yield values should be considered. Thus, the identification of genotypes with wide adaptation would be possible. Therefore, genotypes NS-525, NS-589, and J-103 stand out for their stability and yield above the general average, so we can consider them widely adaptable and recommend them for all investigated localities.
In such cases, when the environments are location-year combinations, Zobel et al. [
83] points out that a site suitable for prediction and recommendation is the one whose interaction effects slightly differ from year to year. Also, it is convenient for the environments to be more discriminating, i.e., informative, which corresponds to less stable environments. Except for KG09, all environments showed contributing interactions and discriminating effect, but KG09 and KG10, as well as ZP09 and ZP10, showed variations in interaction effect, which will cause changes in rank and absolute yield from year to year and will make the production recommendation more difficult. Zaječar, as a locality, is, therefore, the most suitable and the most reliable for examining grain yield, both due to the small difference in the interaction effect between its environments and due to the discriminating effect of both environments. The genotypes recommended for production in Zaječar are the following: Jagodinac, Rekord, NS-293, NS-183, NS-519, and J-96.







{kind=link}
{kind=link}
{kind=link}