
Effects of Extreme-Ambient Temperatures in Silver Barb (Barbonymus gonionotus): Metabolic, Hemato-Biochemical Responses, Enzymatic Activity and Gill Histomorphology



Abstract
:1. Introduction
2. Methodology
2.1. Experimental Fish Collection, Transportation and Conditioning
2.2. Experimental Design
2.3. Measurements of Water Quality
2.4. Evaluation of the Parameters of Growth Performance
2.5. Hemato-Biochemical Responses
2.6. Pepsin Enzymatic Activity
2.7. Histological Examination
2.8. Statistical Analysis
3. Results
3.1. Measurements of Water Quality Parameters
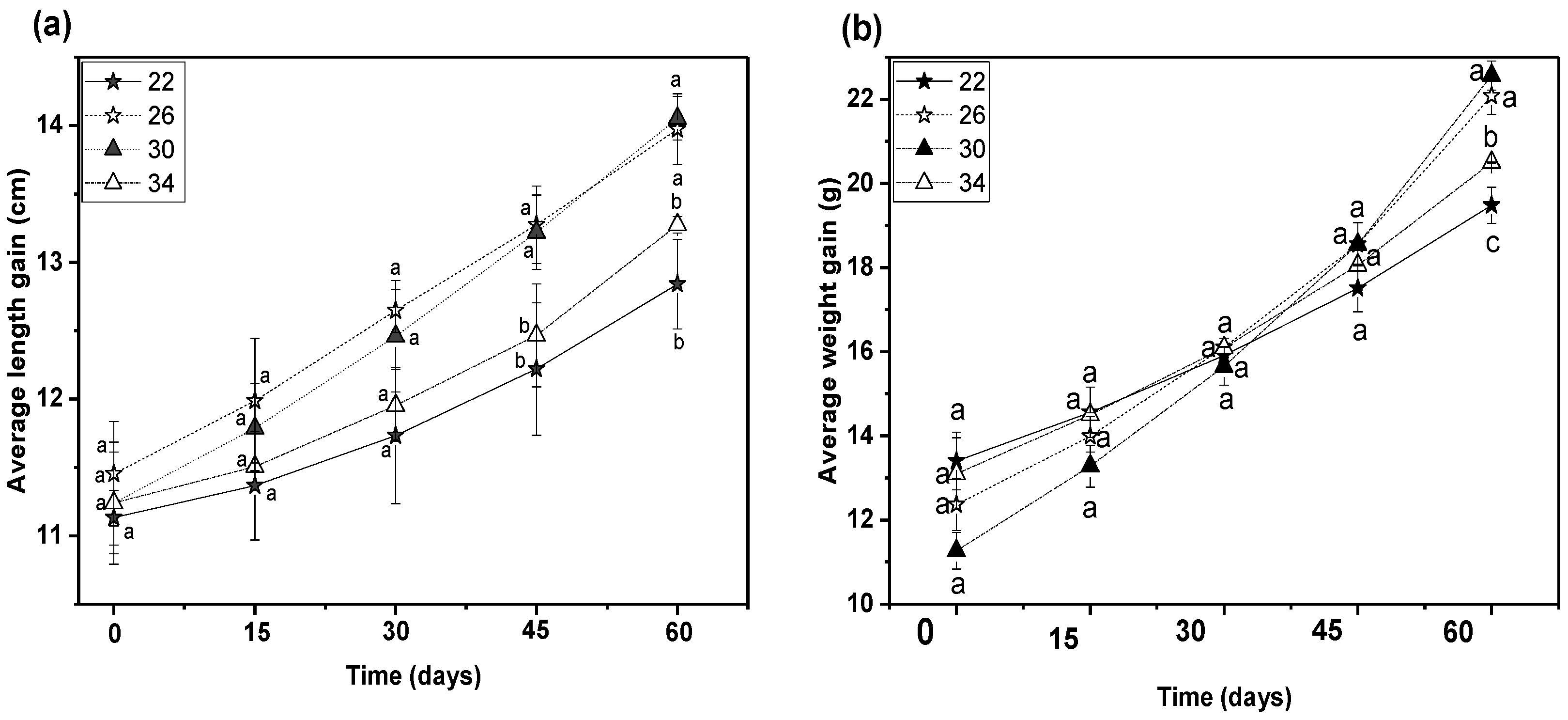
3.2. Growth Performances Study
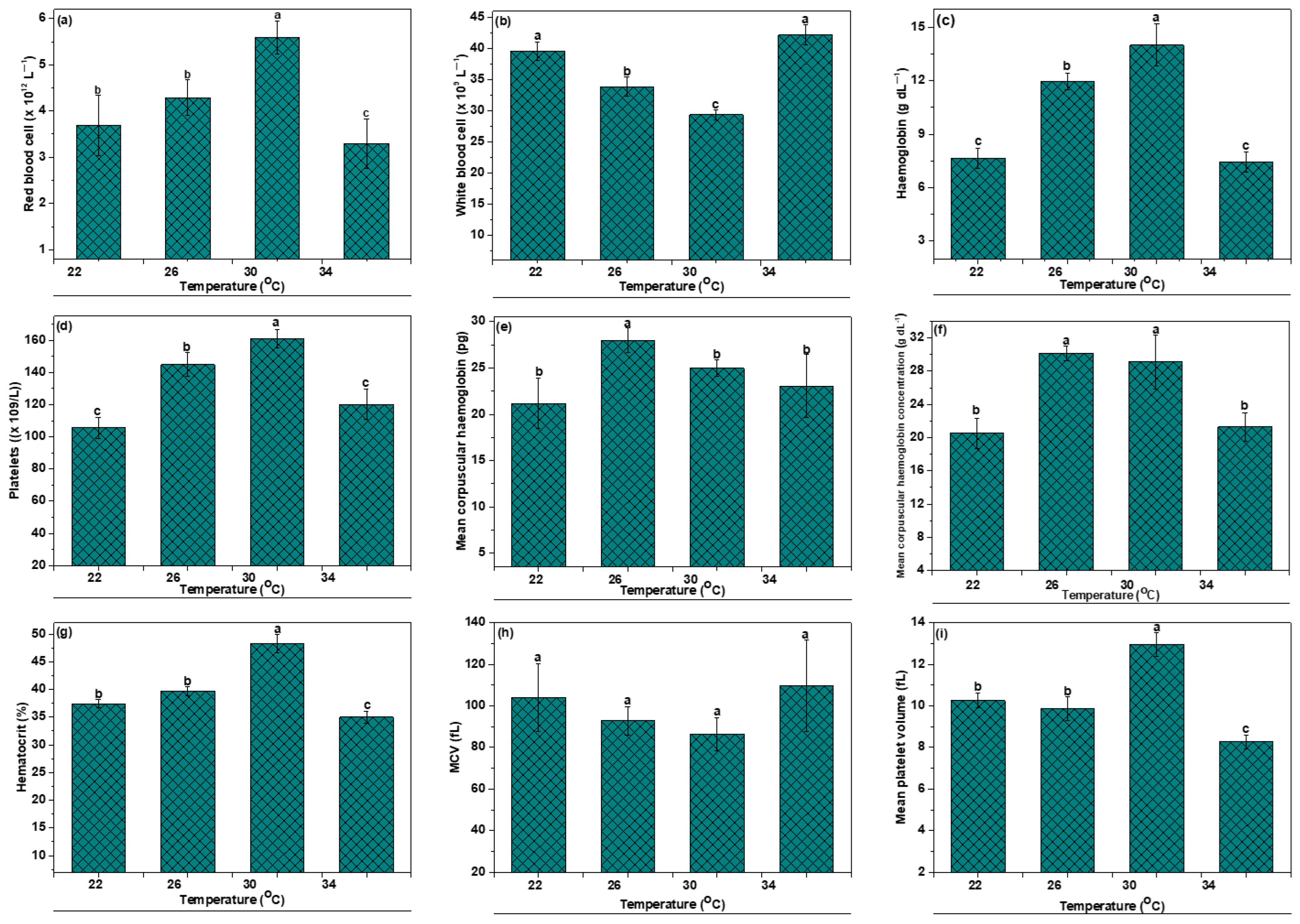
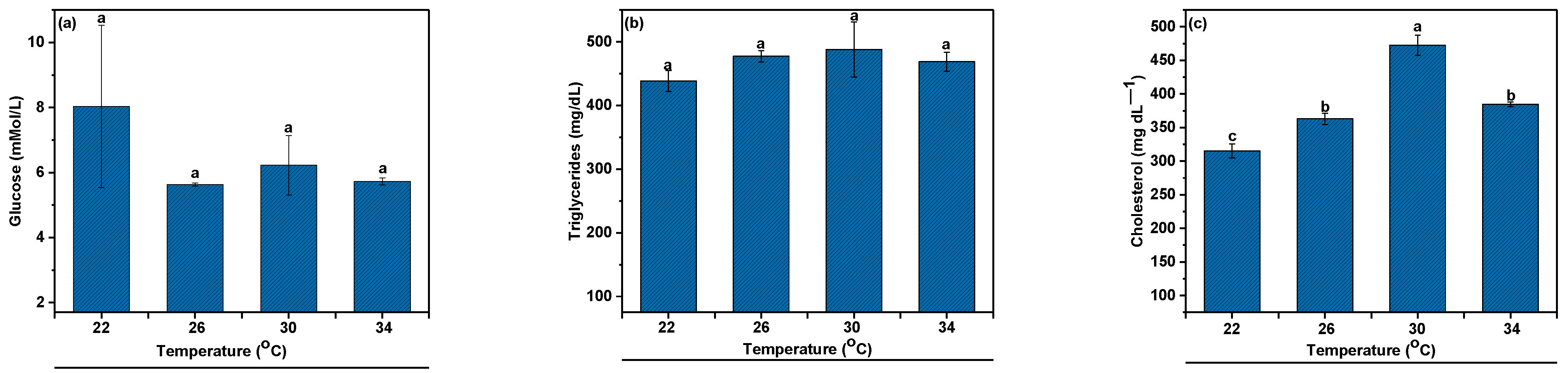
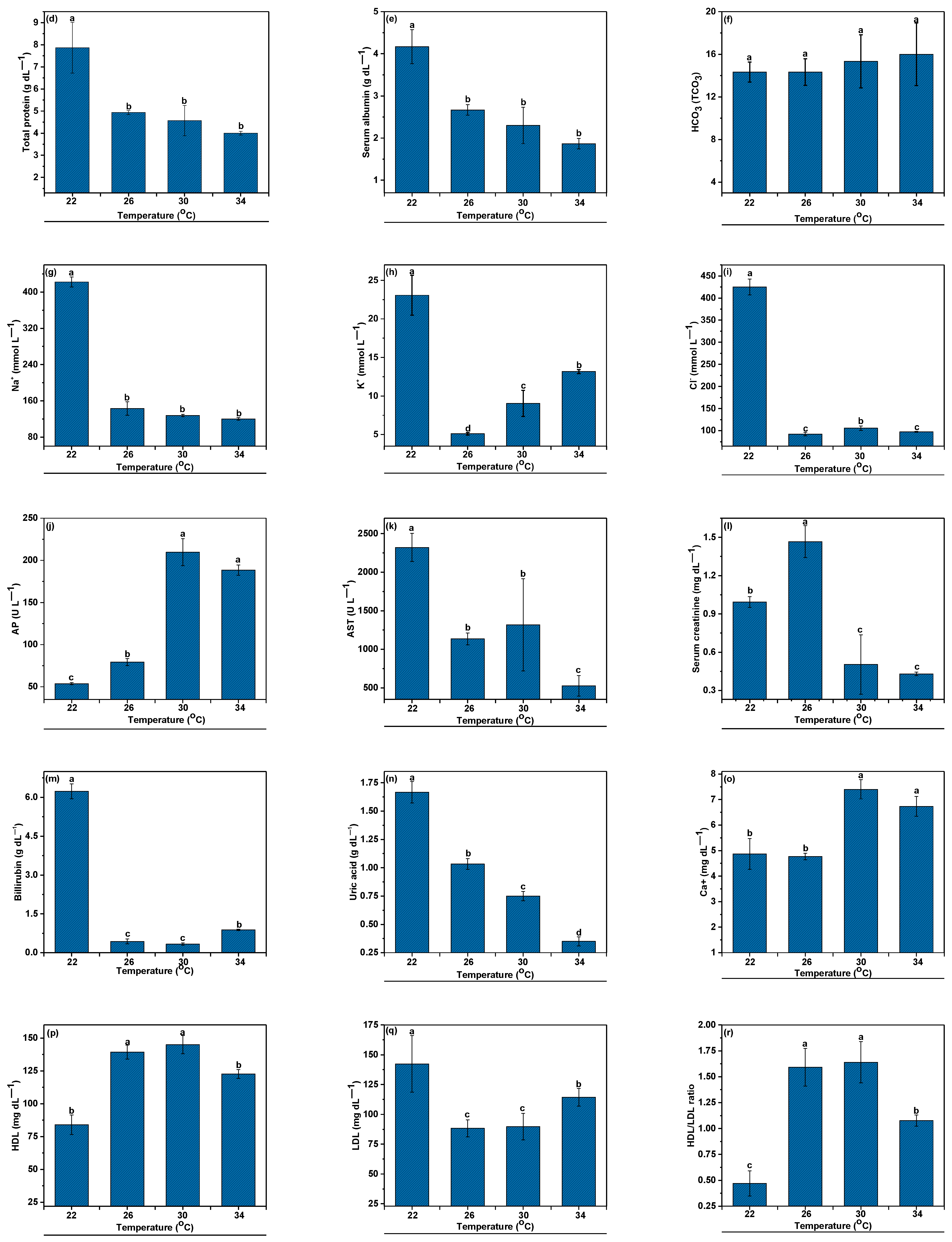
3.3. Hematological Indices
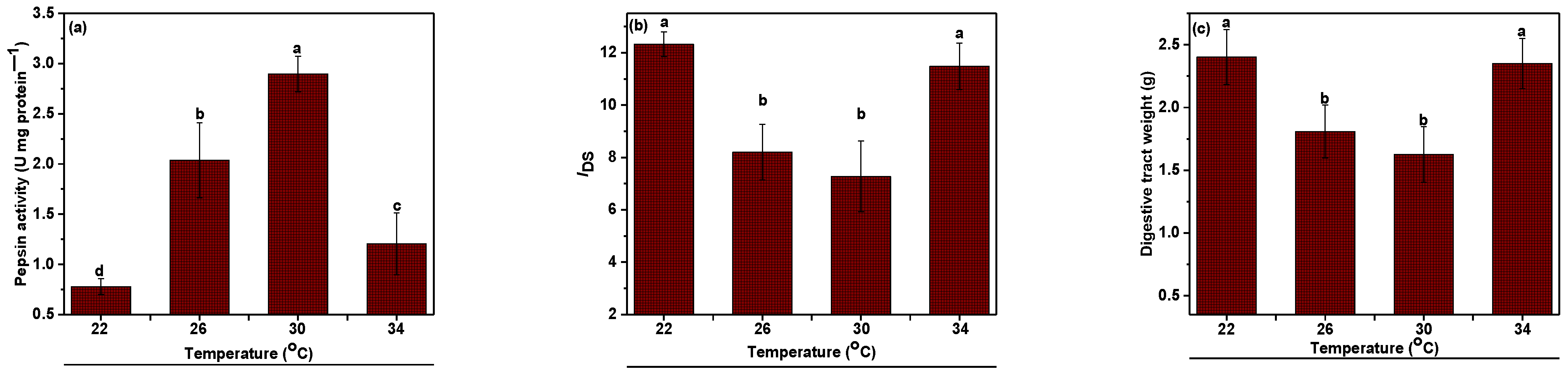
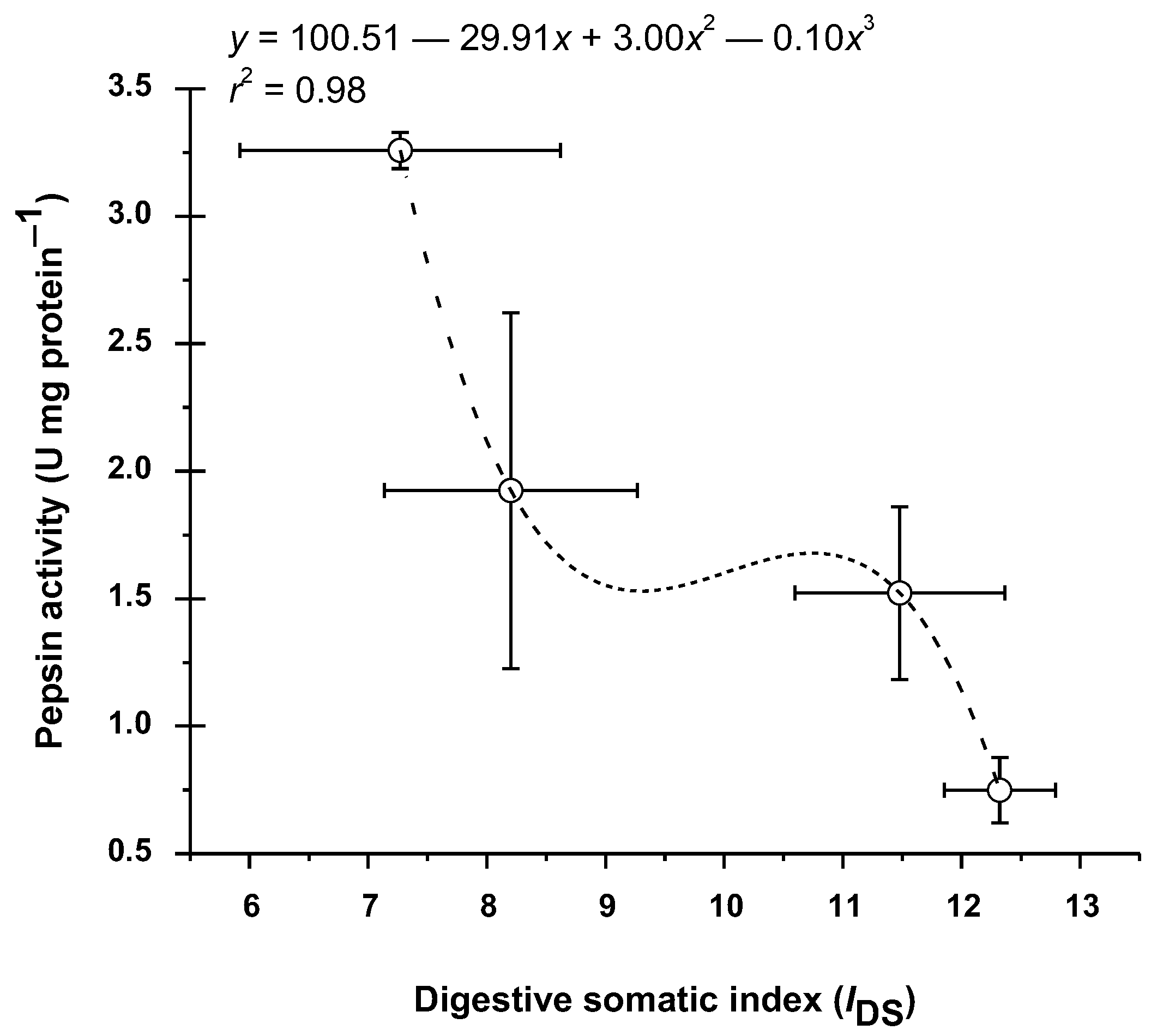
3.4. Pepsin Enzyme Activity
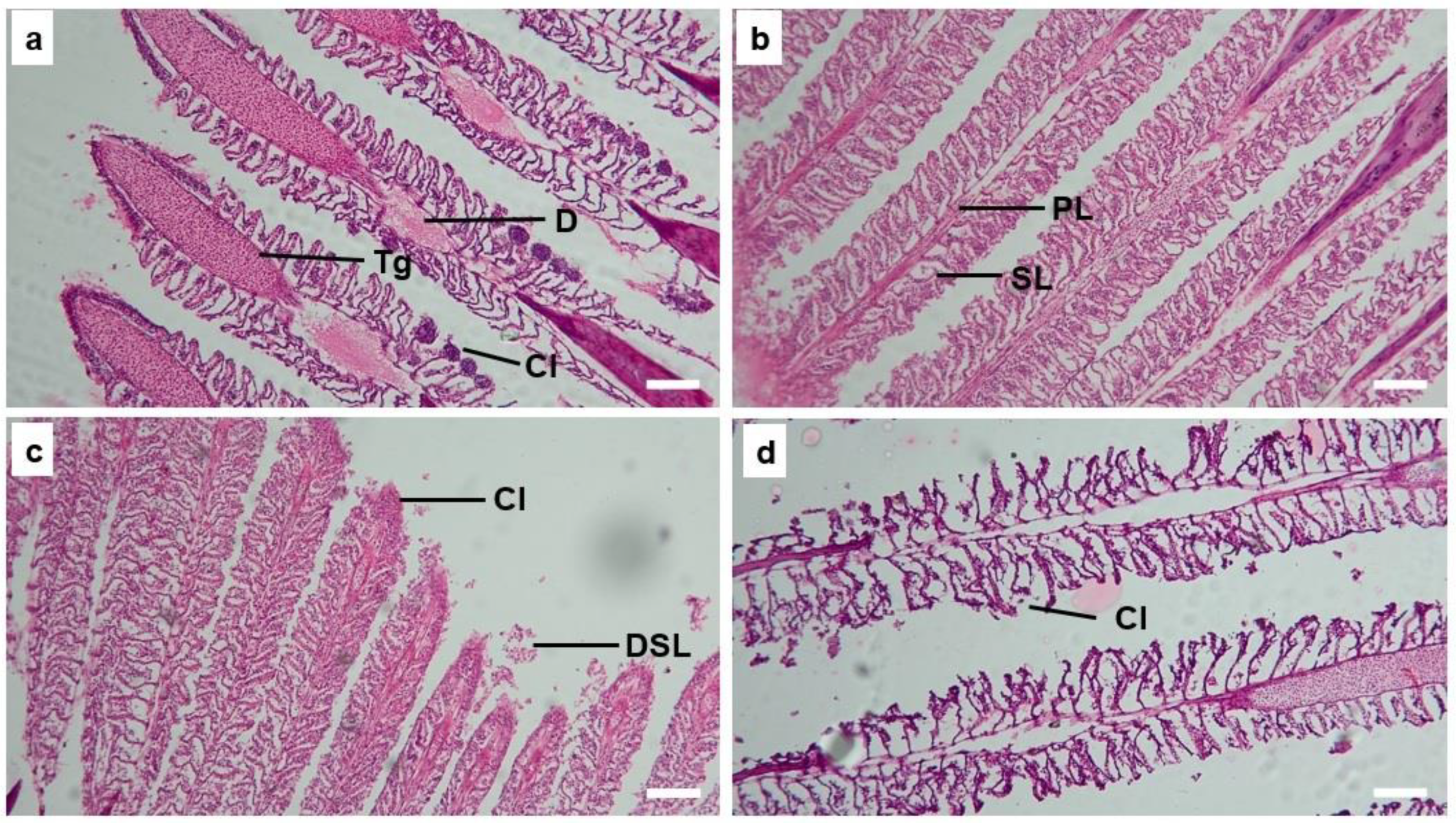
3.5. Gill Histology
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cline, A.J.; Hamilton, S.L.; Logan, C.A. Effects of multiple climate change stressors on gene expression in blue rockfish (Sebastes mystinus). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2020, 239, 110580. [Google Scholar] [CrossRef] [PubMed]
- O’Gorman, E.J.; Ólafsson, Ó.P.; Demars, B.O.; Friberg, N.; Guðbergsson, G.; Hannesdóttir, E.R.; Jackson, M.C.; Johansson, L.S.; McLaughlin, Ó.B.; Ólafsson, J.S.; et al. Temperature effects on fish production across a natural thermal gradient. Glob. Chang. Biol. 2016, 22, 3206–3220. [Google Scholar] [CrossRef] [PubMed]
- Jensen, L.B.; Boltana, S.; Obach, A.; McGurk, C.; Waagbø, R.; MacKenzie, S. Investigating the underlying mechanisms of temperature-related skin diseases in Atlantic salmon, Salmo salar L., as measured by quantitative histology, skin transcriptomics and composition. J. Fish Dis. 2015, 38, 977–992. [Google Scholar] [CrossRef] [PubMed]
- Pfahl, S.; O’Gorman, P.A.; Fischer, E.M. Understanding the regional pattern of projected future changes in extreme precipitation. Nat. Clim. Chang. 2017, 7, 423–427. [Google Scholar] [CrossRef]
- Webster, P.J. Dynamics of the Tropical Atmosphere and Oceans; John Wiley & Sons: Hoboken, NJ, USA, 2020. [Google Scholar]
- Islam, S.M.; Sultana, R.; Imran, M.; Jannat, M.F.T.; Ashaf-Ud-Doulah, M.; Rohani, M.F.; Brown, C.; Shahjahan, M. Elevated temperature affects growth and hemato-biochemical parameters, inducing morphological abnormalities of erythrocytes in Nile tilapia Oreochromis niloticus. Aquac. Res. 2020, 51, 4361–4371. [Google Scholar] [CrossRef]
- Yilmaz, H.A.; Turkmen, S.; Kumlu, M.; Eroldogan, O.T.; Perker, N. Alteration of growth and temperature tolerance of European sea bass (Dicentrarchus labrax linnaeus 1758) in different temperature and salinity combinations. Turk. J. Fish. Aquat. Sci. 2020, 20, 331–340. [Google Scholar] [CrossRef]
- Nadermann, N.; Seward, R.K.; Volkoff, H. Effects of potential climate change-induced environmental modifications on food intake and the expression of appetite regulators in goldfish. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2019, 235, 138–147. [Google Scholar] [CrossRef]
- Burgerhout, E.; Lokman, P.M.; van den Thillart, G.E.; Dirks, R.P. The time-keeping hormone melatonin: A possible key cue for puberty in freshwater eels (Anguilla spp.). Rev. Fish Biol. Fish. 2019, 29, 1–21. [Google Scholar] [CrossRef]
- Cheng, S.Y.; Chen, C.S.; Chen, J.C. Salinity and temperature tolerance of brown-marbled grouper Epinephelus fuscoguttatus. Fish Physiol. Biochem. 2013, 39, 277–286. [Google Scholar] [CrossRef]
- Shahjahan, M.; Kitahashi, T.; Ando, H. Temperature affects sexual maturation through the control of kisspeptin, kisspeptin receptor, GnRH and GTH subunit gene expression in the grass puffer during the spawning season. Gen. Comp. Endocrinol. 2017, 243, 138–145. [Google Scholar] [CrossRef]
- Slater, M.; D’Abramo, L.; Engle, C.R. Aquaculture research priorities for the next decade: A global perspective. J. World Aquac. Soc. 2018, 49, 3–6. [Google Scholar] [CrossRef]
- Mazumder, S.K.; Das, S.K.; Rahim, S.M.; Abd Ghaffar, M. Temperature and diet effect on the pepsin enzyme activities, digestive somatic index and relative gut length of Malabar blood snapper (Lutjanus malabaricus Bloch & Schneider, 1801). Aquac. Rep. 2018, 9, 1–9. [Google Scholar]
- Mazumder, S.K.; Fivelstad, S.; Ghaffar, M.A.; Das, S.K. Haematological and Biochemical Responses of Juvenile Malabar Blood Snapper (Lutjanus malabaricus Bloch & Schneider, 1801) Exposed to Different Rearing Temperatures and Diets. Sains Malays. 2019, 48, 1790–1810. [Google Scholar]
- Pepin, P. Effect of temperature and size on development, mortality, and survival rates of the pelagic early life history stages of marine fish. Can. J. Fish. Aquat. Sci. 1991, 48, 503–518. [Google Scholar] [CrossRef]
- Fu, K.K.; Fu, C.; Qin, Y.L.; Bai, Y.; Fu, S.J. The thermal acclimation rate varied among physiological functions and temperature regimes in a common cyprinid fish. Aquaculture 2018, 495, 393–401. [Google Scholar] [CrossRef]
- Maulvault, A.L.; Barbosa, V.; Alves, R.; Custódio, A.; Anacleto, P.; Repolho, T.; Ferreira, P.P.; Rosa, R.; Marques, A.; Diniz, M. Ecophysiological responses of juvenile seabass (Dicentrarchus labrax) exposed to increased temperature and dietary methylmercury. Sci. Total Environ. 2017, 586, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.D.; Iwama, G.K. Energy cost of NaCl transport in isolated gills of cutthroat trout. Am. J. Physiol.-Regul. Integrat. Comp. Physiol. 1999, 277, R631–R639. [Google Scholar] [CrossRef]
- Adams, S.M. Status and use of biological indicators for evaluating the effects of stress on fish. [Formerly used by DOE/TIC for titles for which valid CODEN was not available. Now invalid] 92-DEC-15; (Country unknown/Code not available). 1990, 8.
- Ruane, N.M.; Nolan, D.T.; Rotllant, J.; Costelloe, J.; Bonga, S.W. Experimental exposure of rainbow trout Oncorhynchus mykiss (Walbaum) to the infective stages of the sea louse Lepeophtheirus salmonis (Krøyer) influences the physiological response to an acute stressor. Fish Shellfish Immunol. 2000, 10, 451–463. [Google Scholar] [CrossRef]
- Sharmin, S.; Shahjahan, M.; Hossain, M.A.; Haque, M.A.; Rashid, H. Histopathological changes in liver and kidney of common carp exposed to sub-lethal doses of malathion. Pak. J. Zool. 2015, 47, 1495–1498. [Google Scholar]
- Islam, S.M.; Rahman, M.A.; Nahar, S.; Uddin, M.H.; Haque, M.M.; Shahjahan, M. Acute toxicity of an organophosphate insecticide sumithion to striped catfish Pangasianodon hypophthalmus. Toxicol. Rep. 2019, 6, 957–962. [Google Scholar] [CrossRef] [PubMed]
- Jaffer, N.S.; Rabee, A.M.; Al-Chalabi, S.M. Biochemical and hematological parameters and histological alterations in fish Cyprinus carpio L. as biomarkers for water pollution with chlorpyrifos. Hum. Ecol. Risk Assess. Int. J. 2017, 23, 605–616.S. [Google Scholar] [CrossRef]
- Nimet, J.; Neves, M.P.; Viana, N.P.; de Arruda Amorim, J.P.; Delariva, R.L. Histopathological alterations in gills of a fish (Astyanax bifasciatus) in neotropical streams: Negative effects of riparian forest reduction and presence of pesticides. Environ. Monit. Assess. 2020, 192, 58. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Hossain, M.Y.; Ahamed, F.; Fatematuzzhura, S.B.; Abdallah, E.M.; Ohtomi, J. Biodiversity in the Padma distributary of the Ganges River, Northwestern Bangladesh: Recommendations for conservation. World J. Zool. 2012, 7, 328–337. [Google Scholar]
- Rahman, M.Z.; Zaman, M.; Hossain, A.; Akter, S.; Rahman, M.M.; Mamun, M.A.A.; Alam, M.M.M.; Khalil, S.M.I. Ovarian Histology and Histopathology of Olive Barb, Puntius sarana Exposed to Endocrine Disrupting Chemical (17-A Methyl Testosterone) in Laboratory Condition. Aquac. Stud. 2023, 24, AQUAST1177. [Google Scholar] [CrossRef]
- Mondal, D.; Rahman, T.; Paul, S.; Islam, M.; Miah, I. Effects of probiotic supplementation on the growth performance of Thai silver barb (Barbonymus gonionotus) (bleeker, 1850) fry. Fundam. Appl. Agric. 2019, 4, 950–958. [Google Scholar] [CrossRef]
- Mohanta, K.N.; Mohanty, S.N.; Jena, J.K.; Sahu, N.P. Apparent protein, lipid and energy digestibility coefficients of some commonly used feed ingredients in formulated pelleted diets for silver barb, Puntius gonionotus. Aquac. Nutr. 2006, 12, 211–218. [Google Scholar] [CrossRef]
- Mostakim, G.M.; Zahangir, M.M.; Mishu, M.M.; Rahman, M.K.; Islam, M.S. Alteration of blood parameters and histoarchitecture of liver and kidney of silver barb after chronic exposure to quinalphos. J. Toxicol. 2015, 2015, 415984. [Google Scholar] [CrossRef]
- Sadiqul, I.M.; Ferdous, Z.; Nannu, M.T.A.; Mostakim, G.M.; Rahman, M.K. Acute exposure to a quinalphos containing insecticide (convoy) causes genetic damage and nuclear changes in peripheral erythrocytes of silver barb, Barbonymus gonionotus. Environ. Pollut. 2016, 219, 949–956. [Google Scholar] [CrossRef]
- Nayak, M.; Saha, A.; Pradhan, A.; Samanta, M.; Giri, S.S. Dietary fish oil replacement by linseed oil: Effect on growth, nutrient utilization, tissue fatty acid composition and desaturase gene expression in silver barb (Puntius gonionotus) fingerlings. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2017, 205, 1–12. [Google Scholar] [CrossRef]
- Jahan, A.; Nipa, T.T.; Islam, S.M.; Uddin, M.H.; Islam, M.S.; Shahjahan, M. Striped catfish (Pangasianodon hypophthalmus) could be suitable for coastal aquaculture. J. Appl. Ichthyol. 2019, 35, 994–1003. [Google Scholar] [CrossRef]
- Wangsongsak, A.; Utarnpongsa, S.; Kruatrachue, M.; Ponglikitmongkol, M.; Pokethitiyook, P.; Sumranwanich, T. Alterations of organ histopathology and metallolhionein mRNA expression in silver barb, Puntius gonionotus during subchronic cadmium exposure. J. Environ. Sci. 2007, 19, 1341–1348. [Google Scholar] [CrossRef] [PubMed]
- Romano, N.; Ashikin, M.; Teh, J.C.; Syukri, F.; Karami, A. Effects of pristine polyvinyl chloride fragments on whole body histology and protease activity in silver barb Barbodes gonionotus fry. Environ. Pollut. 2018, 237, 1106–1111. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.L.; Altindag, A. Haematological parameters of tench (Tinca tinca L., 1758) on acute and chronic exposures of lethal and sublethal treatments of cadmium. Fresenius Environ. Bull. 2004, 13, 1477–1481. [Google Scholar]
- Blaxhall, P.C.; Daisley, K.W. Routine haematological methods for use with fish blood. J. Fish Biol. 1973, 5, 771–781. [Google Scholar] [CrossRef]
- Shah, S.L.; Altindağ, A. Alterations in the immunological parameters of Tench (Tinca tinca L. 1758) after acute and chronic exposure to lethal and sublethal treatments with mercury, cadmium and lead. Turk. J. Vet. Anim. Sci. 2005, 29, 1163–1168. [Google Scholar]
- Wright, G.; Sanderson, J.M. Brain damage and mortality in dogs following pulsatile and non-pulsatile blood flows in extracorporeal circulation. Thorax 1972, 27, 738–747. [Google Scholar] [CrossRef]
- Ye, H.; Xu, M.; Liu, Q.; Sun, Z.; Zou, C.; Chen, L.; Su, N.; Ye, C. Effects of replacing fish meal with soybean meal on growth performance, feed utilization and physiological status of juvenile obscure puffer, Takifugu obscurus. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 216, 75–81. [Google Scholar] [CrossRef]
- Abolfathi, M.; Hajimoradloo, A.; Ghorbani, R.; Zamani, A. Compensatory growth in juvenile roach Rutilus caspicus: Effect of starvation and re-feeding on growth and digestive surface area. J. Fish. Biol. 2012, 81, 1880–1890. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Natalia, Y.; Hashim, R.; Ali, A.; Chong, A. Characterization of digestive enzymes in a carnivorous ornamental fish, the Asian bony tongue Scleropages formosus (Osteoglossidae). Aquaculture 2004, 233, 305–320. [Google Scholar] [CrossRef]
- Anson, M.L. The estimation of pepsin, trypsin, papain, and cathepsin with hemoglobin. J. Gen. Physiol. 1938, 22, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Huggett, R.J. Biomarkers: Biochemical, Physiological, and Histological Markers of Anthropogenic Stress; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Mašković, J.M.; Marković, G.; Mašković, P.Z. Analytical and environmental control of water quality of the Bjelica river. Acta Agric. Serbica 2018, 23, 111–120. [Google Scholar] [CrossRef]
- Schreck, C.B.; Tort, L. The concept of stress in fish. In Fish Physiology; Academic Press: Cambridge, MA, USA, 2016; Volume 35, pp. 1–34. [Google Scholar]
- Beitinger, T.L.; Bennett, W.A. Quantification of the role of acclimation temperature in temperature tolerance of fishes. Environ. Biol. Fish. 2000, 58, 277–288. [Google Scholar] [CrossRef]
- Mazumder, S.K.; Ghaffar, M.A.; Das, S.K. Exploring the suitable temperature and diet for growth and gastric emptying time of juvenile Malabar Blood Snapper (Lutjanus malabaricus Bloch & Schneider, 1801). Thalass. Int. J. Mar. Sci. 2019, 35, 29–41. [Google Scholar]
- Jun, Q.; Pao, X.; Haizhen, W.; Ruiwei, L.; Hui, W. Combined effect of temperature, salinity and density on the growth and feed utilization of Nile tilapia juveniles (Oreochromis niloticus). Aquac. Res. 2012, 43, 1344–1356. [Google Scholar] [CrossRef]
- Handeland, S.O.; Imsland, A.K.; Stefansson, S.O. The effect of temperature and fish size on growth, feed intake, food conversion efficiency and stomach evacuation rate of Atlantic salmon post-smolts. Aquaculture 2008, 283, 36–42. [Google Scholar] [CrossRef]
- Mazumder, S.K.; Das, S.K.; Bakar, Y.; Ghaffar, M.A. Effects of temperature and diet on length-weight relationship and condition factor of the juvenile Malabar blood snapper (Lutjanus malabaricus Bloch & Schneider, 1801). J. Zhejiang Univ. Sci. 2016, 17, 580–590. [Google Scholar]
- Katersky, R.S.; Carter, C.G. Growth efficiency of juvenile barramundi, Lates calcarifer, at high temperatures. Aquaculture 2005, 250, 775–780. [Google Scholar] [CrossRef]
- Handeland, S.; Arnesen, A.; Stefansson, S. Seawater adaptation and growth of post-smolt Atlantic salmon (Salmo salar) of wild and farmed strains. Aquaculture 2003, 220, 367–384. [Google Scholar] [CrossRef]
- Dong, Y.; Dong, S. Growth and oxygen consumption of the juvenile sea cucumber Apostichopus japonicus (Selenka) at constant and fluctuating water temperatures. Aquac. Res. 2006, 37, 1327–1333. [Google Scholar] [CrossRef]
- Jobling, M.; Meløy, O.; Dos Santos, J.; Christiansen, B. The compensatory growth response of the Atlantic cod: Effects of nutritional history. Aquac. Int. 1994, 2, 75–90. [Google Scholar] [CrossRef]
- Diana, J.S. Biology and Ecology of Fishes; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 1995. [Google Scholar]
- Ferreira, F.; Santos, M.M.; Reis-Henriques, M.A.; Vieira, M.N.; Monteiro, N.M. The annual cycle of spermatogenesis in Lipophrys pholis (Blenniidae), a recently proposed sentinel species for pollution monitoring. Ichthyol. Res. 2011, 58, 360–365. [Google Scholar] [CrossRef]
- Islam, M.J.; Kunzmann, A.; Thiele, R.; Slater, M.J. Effects of extreme ambient temperature in European seabass, Dicentrarchus labrax acclimated at different salinities: Growth performance, metabolic and molecular stress responses. Sci. Total Environ. 2020, 735, 139371. [Google Scholar] [CrossRef] [PubMed]
- Shahjahan, M.; Uddin, M.H.; Bain, V.; Haque, M.M. Increased water temperature altered hemato-biochemical parameters and structure of peripheral erythrocytes in striped catfish Pangasianodon hypophthalmus. Fish Physiol. Biochem. 2018, 44, 1309–1318. [Google Scholar] [CrossRef] [PubMed]
- Rhamdia, P.O.J.; de Moraes, G.M.F.R. Características hematológicas de teleósteos brasileiros. IV. Variáveis do jundiá Rhamdia quelen (Pimelodidae). Ciência Rural 2002, 32, 693–698. [Google Scholar]
- Lermen, C.L.; Lappe, R.; Crestani, M.; Vieira, V.P.; Gioda, C.R.; Schetinger, M.R.C.; Baldisserotto, B.; Moraes, G.; Morsch, V.M. Effect of different temperature regimes on metabolic and blood parameters of silver catfish Rhamdia quelen. Aquaculture 2004, 239, 497–507. [Google Scholar] [CrossRef]
- Sikorska, J.; Kondera, E.; Kamiński, R.; Ługowska, K.; Witeska, M.; Wolnicki, J. Effect of four rearing water temperatures on some performance parameters of larval and juvenile crucian carp, Carassius carassius, under controlled conditions. Aquac. Res. 2018, 49, 3874–3880. [Google Scholar] [CrossRef]
- Carvalho, C.S.; Fernandes, M.N. Effect of temperature on copper toxicity and hematological responses in the neotropical fish Prochilodus scrofa at low and high pH. Aquaculture 2006, 251, 109–117. [Google Scholar] [CrossRef]
- Hedayati, A.; Tarkhani, R. Hematological and gill histopathological changes in iridescent shark, Pangasius hypophthalmus (Sauvage, 1878) exposed to sublethal diazinon and deltamethrin concentrations. Fish Physiol. Biochem. 2014, 40, 715–720. [Google Scholar] [CrossRef]
- Cech, J.J.; Brauner, C.J. Techniques in whole animal respiratory physiology. Encycl. Fish Physiol. Genome Environ. 2011, 2, 846–853. [Google Scholar]
- Francis-Floyd, R.; Watson, C.; Petty, D.; Pouder, D.B. Ammonia in Aquatic Systems; UF/IFAS University of Florida (UF)/Institute of Food and Agricultural Sciences (IFAS), FA: Gainesville, FL, USA, 2009; Volume 16, pp. 1–4. [Google Scholar]
- Roberts, R.J. Fish Pathology; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Satheeshkumar, P.; Ananthan, G.; Kumar, D.S.; Jagadeesan, L. Haematology and biochemical parameters of different feeding behaviour of teleost fishes from Vellar estuary, India. Comp. Clin. Pathol. 2012, 21, 1187–1191. [Google Scholar] [CrossRef]
- Rashmeei, M.; Shekarabi, S.P.H.; Mehrgan, M.S.; Paknejad, H. Stimulatory effect of dietary chasteberry (Vitex agnus-castus) extract on immunity, some immune-related gene expression, and resistance against Aeromonas hydrophila infection in goldfish (Carassius auratus). Fish Shellfish Immunol. 2020, 107, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Gheytasi, A.; Hosseini Shekarabi, S.P.; Islami, H.R.; Mehrgan, M.S. Feeding rainbow trout, Oncorhynchus mykiss, with lemon essential oil loaded in chitosan nanoparticles: Effect on growth performance, serum hemato-immunological parameters, and body composition. Aquac. Int. 2021, 29, 2207–2221. [Google Scholar] [CrossRef]
- Jawad, L.A.; Al-Mukhtar, M.A.; Ahmed, H.K. The relationship between haematocrit and some biological parameters of the Indian shad, Tenualosa ilisha (Family Clupeidae). Anim. Biodivers. Conserv. 2004, 27, 47–52. [Google Scholar]
- Tavares-Dias, M.; Frascá-Scorvo, C.; Novato, P.; Moraes, F. Hematological characteristics of hybrid Florida red tilapia, Oreochromis urolepis hornorun x O. mossambicus under intensive rearing. In Proceedings of the International Symposium on Tilapia Aquaculture, Rio de Janeiro, Brazil, 3–7 September 2000; pp. 533–541. [Google Scholar]
- Adeyemo, O.K.; Agbede, S.A.; Olaniyan, A.O.; Shoaga, O.A. The haematological response of Clarias gariepinus to changes in acclimation temperature. Afr. J. Biomed. Res. 2003, 6, 105–108. [Google Scholar] [CrossRef]
- Chavin, W.; Young, J.E. Factors in the determination of normal serum glucose levels of goldfish, Carassius auratus L. Comp. Biochem. Phys. 1970, 33, 629–653. [Google Scholar] [CrossRef]
- Yuan, W.; Li, X.; Kangjian, Y. Effect beta-cypermethrin on GPT and GOT activities of crucian serum. Nongye Kexue Yu Jishu 2005, 6, 20–23. [Google Scholar]
- Chen, M.; Zhang, X.; Gao, T.; Chen, C. Effects of temperature, pH and NaCl on protease activity in digestive tract of young turbot, Scophthalmus maximus. Chin. J. Oceanol. Limnol. 2006, 24, 300–306. [Google Scholar]
- Munilla-Morán, R.; Saborido-Rey, F. Digestive enzymes in marine species. I. Proteinase activities in gut from redfish (Sebastes mentella), seabream (Sparus aurata) and turbot (Scophthalmus maximus). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1996, 113, 395–402. [Google Scholar] [CrossRef]
- Reece, S.Y.; Hamel, J.A.; Sung, K.; Jarvi, T.D.; Esswein, A.J.; Pijpers, J.J.; Nocera, D.G. Wireless solar water splitting using silicon-based semiconductors and earth-abundant catalysts. Science 2011, 334, 645–648. [Google Scholar] [CrossRef] [PubMed]
- Lazo, J.P.; Mendoza, R.; Holt, G.J.; Aguilera, C.; Arnold, C.R. Characterization of digestive enzymes during larval development of red drum (Sciaenops ocellatus). Aquaculture 2007, 265, 194–205. [Google Scholar] [CrossRef]
- Xiong, D.M.; Xie, C.X.; Zhang, H.J.; Liu, H.P. Digestive enzymes along digestive tract of a carnivorous fish Glyptosternum maculatum (Sisoridae, Siluriformes). J. Anim. Physiol. Anim. Nutr. 2011, 95, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.A.; Cossins, A.R. Adaptation of intestinal morphology in the temperature-acclimated carp, Cyprinus carpio L. Cell Tissue Res. 1988, 251, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Furné, M.; García-Gallego, M.; Hidalgo, M.C.; Morales, A.E.; Domezain, A.; Domezain, J.; Sanz, A. Effect of starvation and refeeding on digestive enzyme activities in sturgeon (Acipenser naccarii) and trout (Oncorhynchus mykiss). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2008, 149, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Krogdahl, Å.; Bakke-McKellep, A.M. Fasting and refeeding cause rapid changes in intestinal tissue mass and digestive enzyme capacities of Atlantic salmon (Salmo salar L.). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2005, 141, 450–460. [Google Scholar] [CrossRef] [PubMed]
- Storebakken, T.; Shearer, K.D.; Refstie, S.; Lagocki, S.; McCool, J. Interactions between salinity, dietary carbohydrate source and carbohydrate concentration on the digestibility of macronutrients and energy in rainbow trout (Oncorhynchus mykiss). Aquaculture 1998, 163, 347–359. [Google Scholar] [CrossRef]
- Bélanger, F.; Blier, P.U.; Dutil, J.D. Digestive capacity and compensatory growth in Atlantic cod (Gadus morhua). Fish Physiol. Biochem. 2002, 26, 121–128. [Google Scholar] [CrossRef]
- Fu, X.; Xue, C.; Miao, B.; Li, Z.; Gao, X.; Hirata, T. Distribution and seasonal activity variation of proteases in digestive tract of sea cucumber Stichopus japonicus. Fish. Sci. 2006, 72, 1130–1132. [Google Scholar] [CrossRef]
- Gao, F.; Yang, H.; Xu, Q.; Wang, F.; Liu, G. Effect of water temperature on digestive enzyme activity and gut mass in sea cucumber Apostichopus japonicus (Selenka), with special reference to aestivation. Chin. J. Oceanol. Limnol. 2009, 27, 714–722. [Google Scholar] [CrossRef]
- Islam, M.R.; Hossain, M.A.; Afrose, F.; Roy, N.C.; Iqbal, M.M. Effect of temperature on the growth performance, haematological properties and histomorphology of gill, intestine and liver tissues in juvenile butter catfish Ompok bimaculatus. Aquac. Fish Fish. 2022, 2, 277–286. [Google Scholar] [CrossRef]
- Maftuch, M.; Sanoesi, E.; Farichin, I.; Saputra, B.A.; Ramdhani, L.; Hidayati, S.; Fitriyah, N.; Prihanto, A.A. Histopathology of gill, muscle, intestine, kidney, and liver on Myxobolus sp.-infected Koi carp (Cyprinus carpio). J. Parasit. Dis. 2018, 42, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Amir, F.; Muchlisin, Z.A.; Nur, F.M.; Fadli, N.; Siti-Azizah, M.N.; Wilkes, M.; Tang, U.; Hasan, B.; Batubara, A.S.; Kocabas, F.K.; et al. Effect of increasing water temperature on the physiology and gill histology of Barramundi, Lates calcarifer (Pisces, Perciformes) fingerlings. Int. Aquat. Res. 2022, 14, 263–273. [Google Scholar]
- Flores-Lopes, F.; Thomaz, A.T. Histopathologic alterations observed in fish gills as a tool in environmental monitoring. Braz. J. Biol. 2011, 71, 179–188. [Google Scholar] [CrossRef]
- Evans, D.H.; Piermarini, P.M.; Choe, K.P. The multifunctional fish gill: Dominant site of gas exchange, osmoregulation, acid-base regulation, and excretion of nitrogenous waste. Physiol. Rev. 2005, 85, 97–177. [Google Scholar] [CrossRef]
- Saber, T.H. Histological adaptation to thermal changes in gills of common carp fishes Cyprinus carpio L. Rafidain J. Sci. 2011, 22, 46–55. [Google Scholar] [CrossRef]
- Zaman, M.; Khalil, S.M.I.; Rahman, M.Z.; Hossain, A.; Al Mamun, M.A.; Rahman, M.M.; Alam, M.M. Evaluation of Histopathological Alterations in the Liver and Kidney of Olive Barb (Puntius sarana, Hamilton 1822) as an Indicator of the Surma River’s Pollution. Aquat. Sci. Eng. 2023, 38, 81–88. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | 22 °C | 26 °C | 30 °C | 34 °C |
|---|---|---|---|---|
| Temperature (°C) | 22 ± 0.84 | 26 ± 0.62 | 30 ± 0.38 | 34 ± 0.61 |
| * TH (mg L−1) | 123.33 ± 17.33 | 110.36 ± 24.36 | 118.29 ± 21.37 | 112.16 ± 16.21 |
| * NH3-N (mg L−1) | 0.24 ± 0.054 | 0.16 ± 0.018 | 0.31 ± 0.024 | 0.19 ± 0.031 |
| pH | 6.91 ± 0.06 | 7.51 ± 0.24 | 7.66 ± 0.63 | 7.81 ± 0.96 |
| Variables | Temperature (°C) | |||
|---|---|---|---|---|
| 22 | 26 | 30 | 34 | |
| Initial total length (cm) | 11.13 ± 1.00 a | 11.45 ± 1.38 a | 11.24 ± 1.24 a | 11.24 ± 1.14 a |
| Final total length (cm) | 12.84 ± 1.23 a | 13.97 ± 1.93 a | 14.05 ± 1.66 a | 13.27 ± 1.60 a |
| Initial body weight (g) | 13.40 ± 3.68 a | 12.37 ± 4.62 a | 11.27 ± 2.43 a | 13.10 ± 3.86 a |
| Final body weight (g) | 19.48 ± 4.96 a | 22.08 ± 6.44 b | 22.57 ± 4.35 b | 20.50 ± 5.01 a |
| Total length gain (cm) | 1.71 ± 0.31 b | 2.52 ± 0.61 a | 2.81 ± 0.46 a | 2.03 ± 0.49 b |
| Body weight gain (g) | 6.08 ± 1.42 c | 9.71 ± 1.93 a | 11.30 ± 1.96 a | 7.40 ± 1.17 b |
| Food conversion ratio | 1.83 ± 0.05 a | 1.51 ± 0.03 c | 1.42 ± 0.04 d | 1.75 ± 0.02 b |
| Food conversion efficiency | 0.55 ± 0.01 c | 0.66 ± 0.02 b | 0.70 ± 0.01 a | 0.57 ± 0.02 d |
| Condition factor | 0.90 ± 0.09 a | 0.81 ± 0.11 a | 0.83 ± 0.14 a | 0.87 ± 0.11 a |
| Specific growth rate (% day−1) | 0.63 ± 0.07 d | 1.01 ± 0.13 b | 1.17 ± 0.06 a | 0.77 ± 0.10 c |
| Relative growth rate (%) | 59.34 ± 10.05 c | 83.41 ± 14.23 b | 101.32 ± 6.99 a | 45.99 ± 5.81 d |
| Daily growth rate (%) | 40.53 ± 9.49 d | 97.13 ± 19.26 b | 113.00 ± 19.59 a | 74.00 ± 11.73 c |
| Survival rate (%) | 100 | 100 | 100 | 100 |
| Type of Abnormality | Temperature (°C) | |||
|---|---|---|---|---|
| 22 | 26 | 30 | 34 | |
| Clubbing | +++ | ++ | ++ | +++ |
| Hypertrophy | ++ | 0 | + | + |
| Hyperplasia | + | 0 | + | + |
| Telangiectasis | +++ | ++ | ++ | ++ |
| Degeneration or detachment of SL | + | + | ++ | + |
| Inflammatory cell infiltration | +++ | ++ | + | ++ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazumder, S.K.; Debi, S.; Das, S.K.; Salam, M.A.; Alam, M.S.; Rahman, M.L.; Mamun, M.A.A.; Ibrahim Khalil, S.M.; Pandit, D. Effects of Extreme-Ambient Temperatures in Silver Barb (Barbonymus gonionotus): Metabolic, Hemato-Biochemical Responses, Enzymatic Activity and Gill Histomorphology. Water 2024, 16, 292. https://doi.org/10.3390/w16020292
Mazumder SK, Debi S, Das SK, Salam MA, Alam MS, Rahman ML, Mamun MAA, Ibrahim Khalil SM, Pandit D. Effects of Extreme-Ambient Temperatures in Silver Barb (Barbonymus gonionotus): Metabolic, Hemato-Biochemical Responses, Enzymatic Activity and Gill Histomorphology. Water. 2024; 16(2):292. https://doi.org/10.3390/w16020292
Chicago/Turabian StyleMazumder, Sabuj Kanti, Sutapa Debi, Simon Kumar Das, Mohammad Abdus Salam, Md. Shahanoor Alam, Mohammad Lutfar Rahman, Md. Abdullah Al Mamun, Sarker Mohammed Ibrahim Khalil, and Debasish Pandit. 2024. "Effects of Extreme-Ambient Temperatures in Silver Barb (Barbonymus gonionotus): Metabolic, Hemato-Biochemical Responses, Enzymatic Activity and Gill Histomorphology" Water 16, no. 2: 292. https://doi.org/10.3390/w16020292