
Three New Species of Russulaceae (Russulales, Basidiomycota) from Southern China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Collections and Morphological Analyses
2.2. DNA Extraction, PCR Amplification, and Sequencing
2.3. Alignment and Phylogenetic Analyses
3. Results
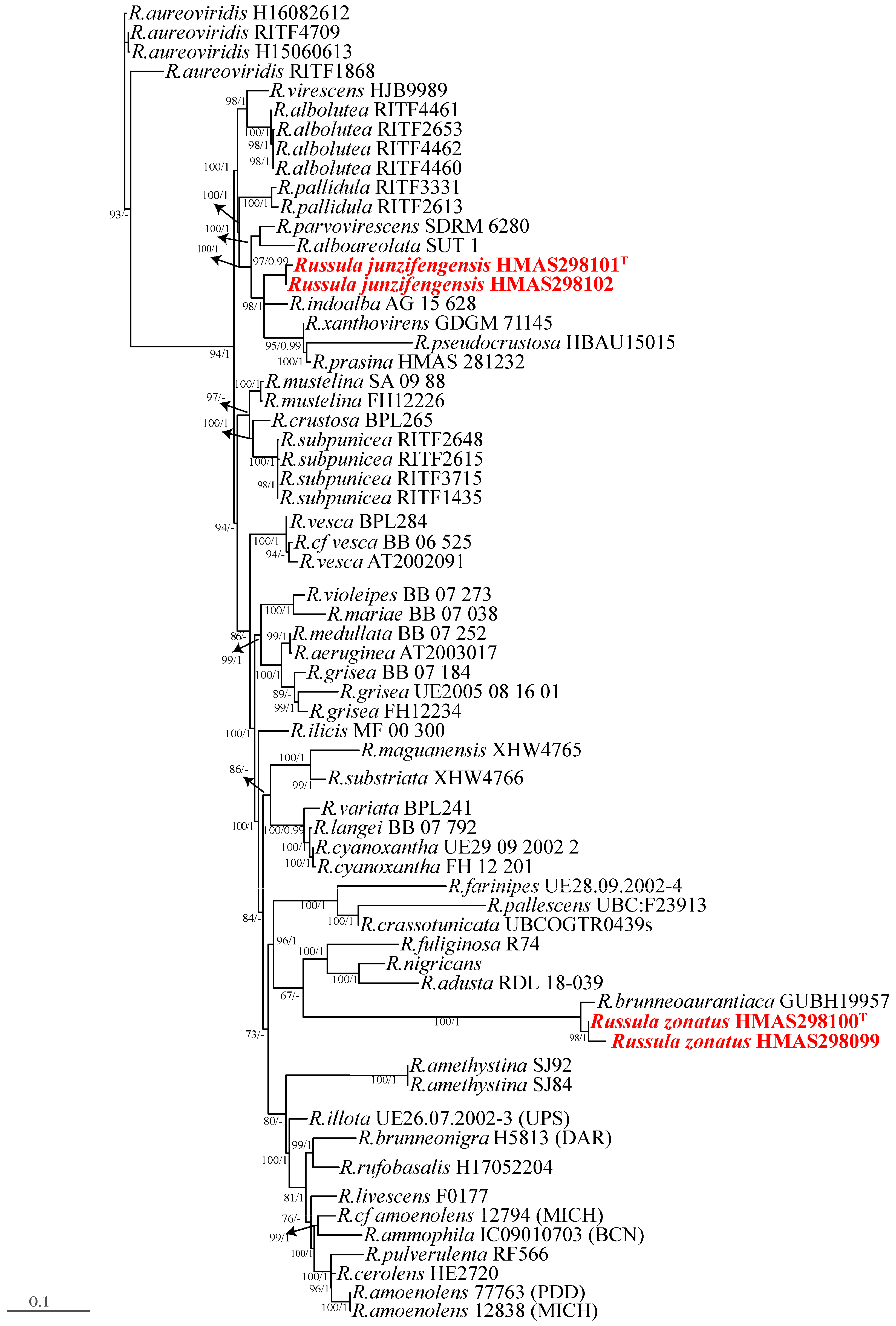
3.1. Phylogenetic Analyses
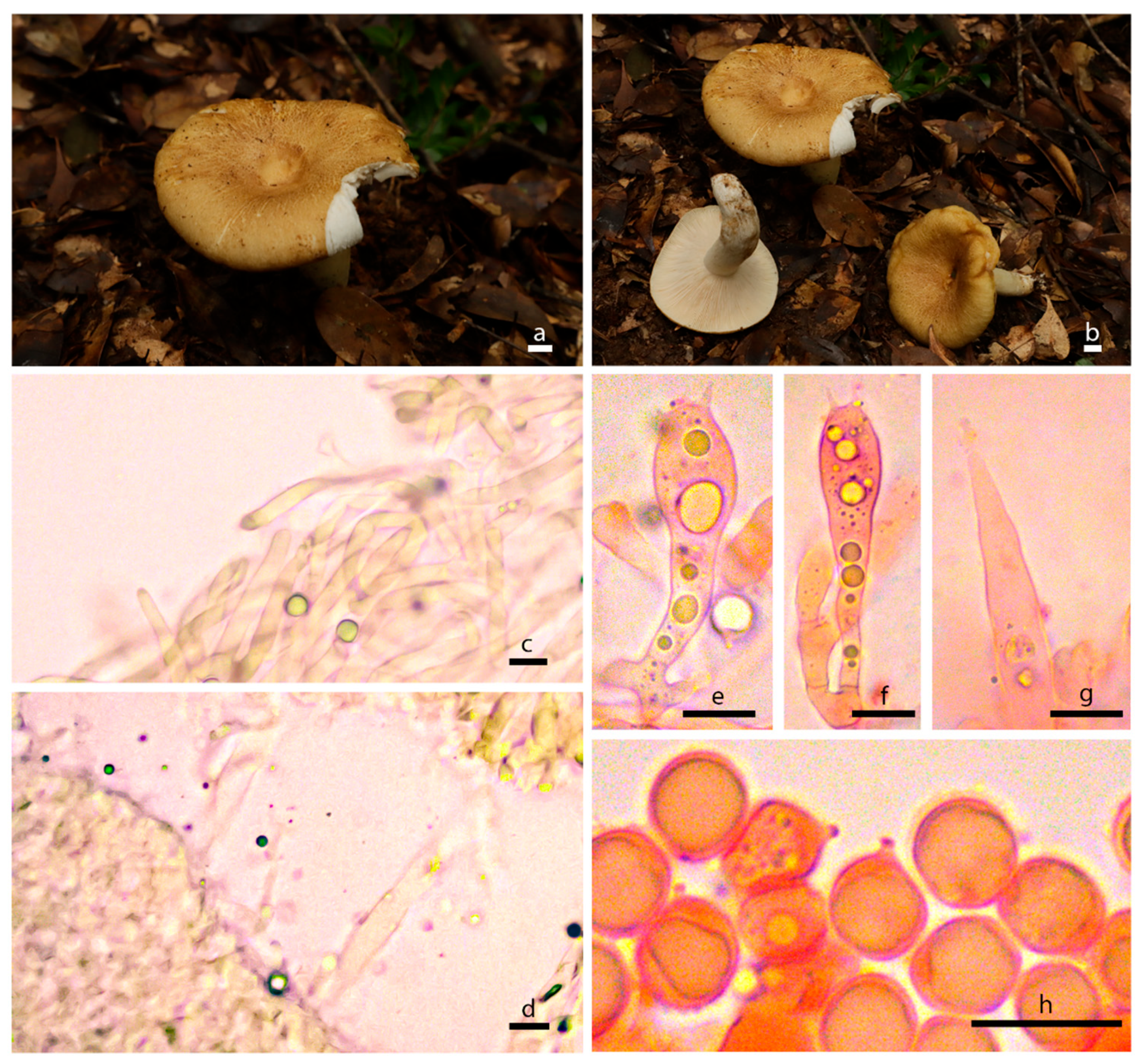
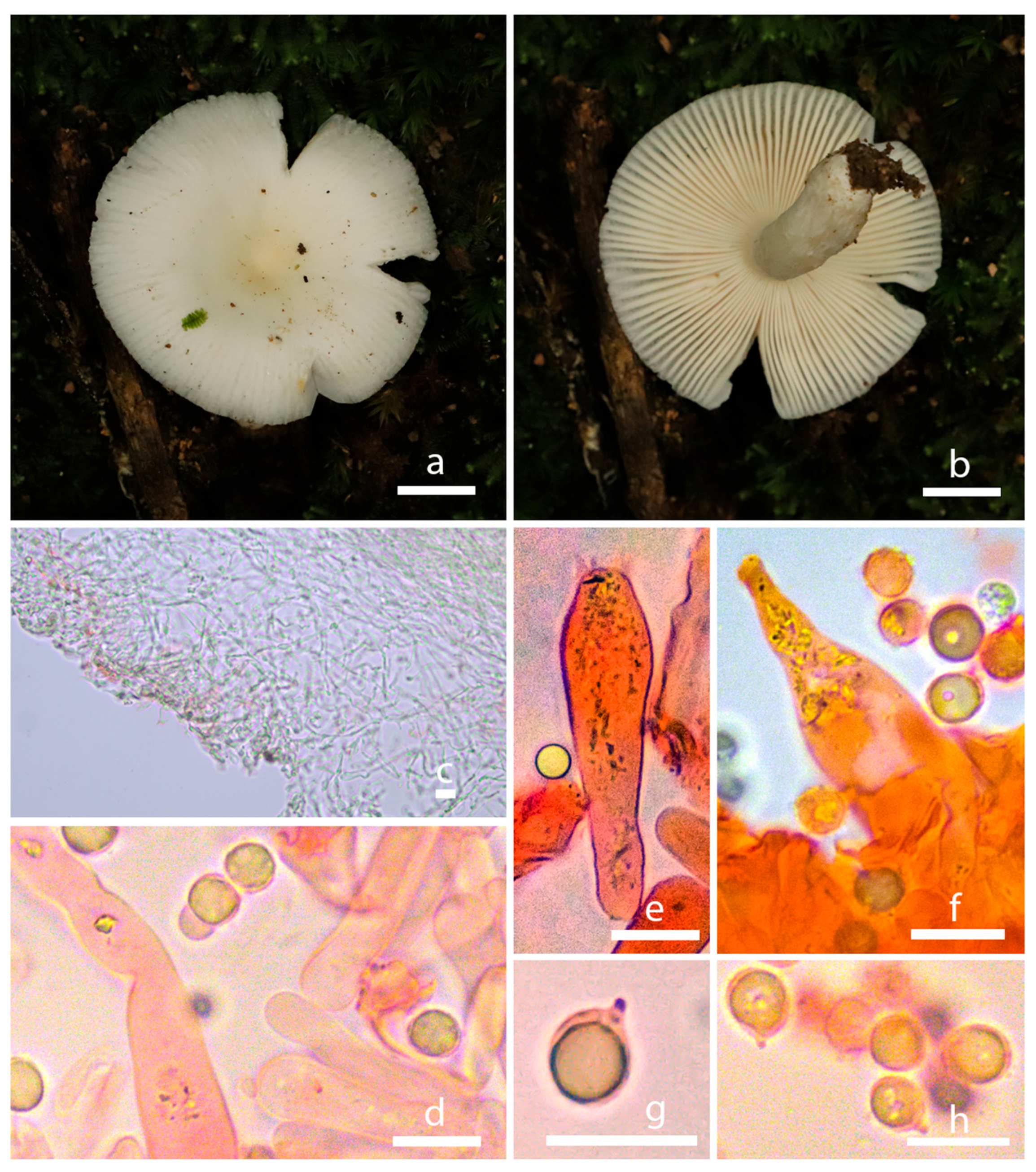
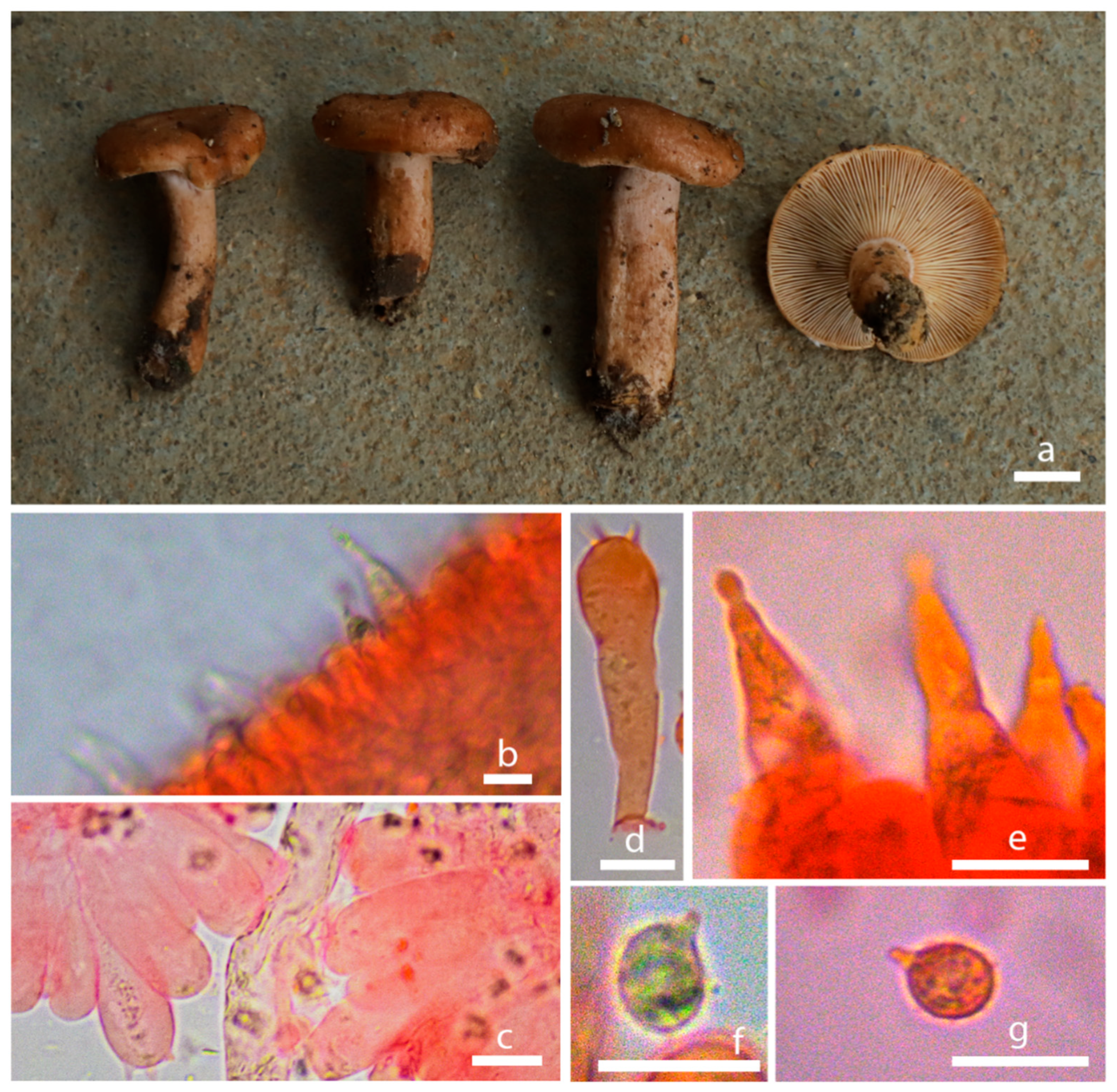
3.2. Taxonomy
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Looney, B.P.; Meidl, P.; Piatek, M.J.; Miettinen, O.; Martin, F.M.; Matheny, P.B.; Labbé, J.L. Russulaceae: A new genomic dataset to study ecosystem function and evolutionary diversification of ectomycorrhizal fungi with their tree associates. New Phytol. 2018, 218, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.M.; Li, Y.K.; Liang, J.F.; Wu, J.R. Russula brunneovinacea sp. nov., from Northeastern China. Mycotaxon 2018, 132, 789–797. [Google Scholar] [CrossRef]
- Li, G.J.; Zhao, D.; Li, S.F.; Wen, H.A. Russula chiui and R. pseudopectinatoides, two new species from Southwestern China supported by morphological and molecular evidence. Mycol. Prog. 2015, 14, 33. [Google Scholar] [CrossRef]
- Taylor, A.F.S.; Alexander, I.J. Ectomycorrhizal synthesis with an isolate of Russula aeruginea. Mycol. Res. 1989, 92, 103–107. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, Y.; Mi, F.; Tang, X.; He, X.; Cao, Y.; Liu, C.; Yang, D.; Dong, J.; Zhang, K.; et al. Recent advances in population genetics of ectomycorrhizal mushrooms Russula spp. Mycology 2015, 6, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.K. Biologically active substances from mushrooms in Yunnan, China. Heterocycles 2002, 57, 157–167. [Google Scholar] [CrossRef]
- Singh, R.S.; Walia, A.K.; Kennedy, J.F. Mushroom lectins in biomedical research and development. Int. J. Biol. Macromol. 2020, 151, 1340–1350. [Google Scholar] [CrossRef]
- Ali, A.; Guo, D.; Mahar, A.; Wang, P.; Shen, F.; Li, R.H.; Zhang, Z.Q. Mycoremediation of potentially toxic trace elements—A biological tool for soil cleanup: A review. Pedosphere 2017, 27, 205–222. [Google Scholar] [CrossRef]
- Chen, F.; Yuan, Y.; Wei, W.; Yu, S.; Zhang, T. Reconstructed Temperature for Yong’an, Fujian, Southeast China: Linkages to the pacific ocean climate variability. Glob. Planet. Chang. 2012, 86–87, 11–19. [Google Scholar] [CrossRef]
- Yin, Y.; Gemmer, M.; Luo, Y.; Wang, Y. Tropical cyclones and heavy rainfall in Fujian province, China. Quat. Int. 2010, 226, 122–128. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, X.; Dimitrov, D.; Pellissier, L.; Borregaard, M.K.; Shrestha, N.; Su, X.; Luo, A.; Zimmermann, N.E.; Rahbek, C.; et al. An updated floristic map of the world. Nat. Commun. 2023, 14, 2990. [Google Scholar] [CrossRef] [PubMed]
- Yue, Y.; Zheng, Z.; Huang, K.; Chevalier, M.; Chase, B.M.; Carré, M.; Ledru, M.P.; Cheddadi, R. A Continuous record of vegetation and climate change over the past 50,000 years in the Fujian province of eastern subtropical China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 365–366, 115–123. [Google Scholar] [CrossRef]
- Wisitrassameewong, K.; Park, M.S.; Lee, H.; Ghosh, A.; Das, K.; Buyck, B.; Looney, B.P.; Caboň, M.; Adamčík, S.; Kim, C.; et al. Taxonomic revision of Russula subsection Amoeninae from South Korea. MycoKeys 2020, 75, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Li, G.J.; Dong, Z.; Li, S.F.; Wen, H.A. Recent research progress of Russula (Russulales, Agaricomycetes): A review. Mycosystema 2015, 34, 821–848. [Google Scholar] [CrossRef]
- Vera, M.; Adamčík, S.; Adamčíková, K.; Hampe, F.; Caboň, M.; Manz, C.; Ovrebo, C.; Piepenbring, M.; Corrales, A. Morphological and genetic diversification of Russula floriformis, sp. nov., along the Isthmus of Panama. Mycologia 2021, 113, 807–827. [Google Scholar] [CrossRef]
- Khatua, S.; Paloi, S.; Acharya, K. An untold story of a novel mushroom from Tribal Cuisine: An Ethno-Medicinal, taxonomic and pharmacological approach. Food Funct. 2021, 12, 4679–4695. [Google Scholar] [CrossRef]
- Panda, M.K.; Das, S.K.; Mohapatra, S.; Debata, P.R.; Tayung, K.; Thatoi, H. Mycochemical composition, bioactivities, and phylogenetic placement of three wild edible Russula species from Northern Odisha, India. Plant Biosyst. 2021, 155, 1041–1055. [Google Scholar] [CrossRef]
- Looney, B.P.; Manz, C.; Matheny, P.B.; Adamčík, S. Systematic revision of the Roseinae clade of Russula, with a focus on eastern North American taxa. Mycologia 2022, 114, 270–302. [Google Scholar] [CrossRef]
- Noffsinger, C.; Cripps, C.L. Systematic analysis of Russula in the North American Rocky Mountain alpine zone. Mycologia 2021, 113, 1278–1315. [Google Scholar] [CrossRef]
- Paloi, S.; Kumla, J.; Karunarathna, S.C.; Lumyong, S.; Suwannarach, N. Taxonomic and phylogenetic evidence reveal two new Russula species (Russulaceae, Russulales) from Northern Thailand. Mycol. Prog. 2023, 22, 72. [Google Scholar] [CrossRef]
- Miller, S.L.; Buyck, B. Molecular phylogeny of the genus Russula in Europe with a comparison of modern infrageneric classifications. Mycol. Res. 2002, 106, 259–276. [Google Scholar] [CrossRef]
- Buyck, B.; Hofstetter, V.; Eberhardt, U.; Verbeken, A.; Kauff, F. Walking the thin line between Russula and Lactarius: The dilemma of Russula subsect. Ochricompactae. Fungal Divers. 2008, 28, 15–40. [Google Scholar]
- Buyck, B.; Hofstetter, V.; Verbeken, A.; Walleyn, R. Proposal to conserve Lactarius Nom. Cons. (Basidiomycota) with a conserved type. Taxon 2010, 59, 295–296. [Google Scholar] [CrossRef]
- Li, G.J.; Liu, T.Z.; Li, S.M.; Zhao, S.Y.; Niu, C.Y.; Liu, Z.Z.; Xie, X.J.; Zhang, X.; Shi, L.Y.; Guo, Y.B.; et al. Four new species of Russula subsection Sardoninae from China. J. Fungi 2023, 9, 199. [Google Scholar] [CrossRef]
- Zhou, H.; Cheng, G.Q.; Wang, Q.T.; Guo, M.J.; Zhuo, L.; Yan, H.F.; Li, G.J.; Hou, C.L. Morphological characteristics and phylogeny reveal six new species in Russula Subgenus Russula (Russulaceae, Russulales) from Yanshan Mountains, North China. J. Fungi 2022, 8, 1283. [Google Scholar] [CrossRef]
- Wang, S.H.; Li, G.J.; Phurbu, D.; He, M.Q.; Zhang, M.Z.; Zhu, X.Y.; Li, J.X.; Zhao, R.L.; Cao, B. Four new species of Russula from the Xizang Autonomous Region and other provinces of China. Mycology 2023, 14, 1–28. [Google Scholar] [CrossRef]
- Li, G.J.; Li, S.M.; Buyck, B.; Zhao, S.Y.; Xie, X.J.; Shi, L.Y.; Deng, C.Y.; Meng, Q.F.; Sun, Q.B.; Yan, J.Q.; et al. Three new Russula Species in Sect. Ingratae (Russulales, Basidiomycota) from Southern China. MycoKeys 2021, 84, 103–139. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Song, J.; Liang, J.; Li, Y. Two new species of Russula Subsect. Virescentinae from Southern China. Mycol. Prog. 2021, 20, 993–1005. [Google Scholar] [CrossRef]
- Li, F.; Deng, Q.L. Three new species of Russula from South China. Mycol. Prog. 2018, 17, 1305–1321. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols; Elsevier: Amsterdam, The Netherlands, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1- sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.J.; Hall, B.D. Body plan evolution of Ascomycetes, as inferred from an RNA polymerase II phylogeny. Proc. Natl. Acad. Sci. USA 2004, 101, 4507–4512. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Song, J.; Chen, Y.; Zhang, J.; Liang, J. Morphological and phylogenetic evidence for two new species of Russula Subg. Heterophyllidia from Guangdong province of China. MycoKeys 2021, 82, 139–157. [Google Scholar] [CrossRef]
- Deng, L.S.; Kang, R.; Zeng, N.K.; Yu, W.J.; Chang, C.; Xu, F.; Deng, W.Q.; Qi, L.L.; Zhou, Y.L.; Fan, Y.G. Two new Inosperma (Inocybaceae) species with unexpected muscarine contents from tropical China. MycoKeys 2021, 85, 87–108. [Google Scholar] [CrossRef]
- Buyck, B. One step closer to unravelling the origin of Russula: Subgenus Glutinosae subg. nov. Mycosphere 2020, 11, 285–304. [Google Scholar] [CrossRef]
- Roy, N.; Beypih, J.; Tanti, B.; Dutta, A.K. Russula brunneoaurantiaca, a novel taxon of Russula subg. Crassotunicata from west Bengal, India, with morpho-molecular analysis and scanning electron microscopy. Microsc. Res. Tech. 2023, 86, 1–7. [Google Scholar] [CrossRef]
- Shi, S.F.; Wang, X.H.; Bau, T. Three new species of Lactarius (Russulaceae, Russulales) from Northeast China. Mycoscience 2018, 59, 206–217. [Google Scholar] [CrossRef]
- Wang, X.H. Seven new species of Lactarius subg. Lactarius (Russulaceae) from Southwestern China. Mycosystema 2017, 36, 1463–1482. [Google Scholar] [CrossRef]
- Lee, H.; Park, M.S.; Jung, P.E.; Eimes, J.A.; Seok, S.J.; Lim, Y.W. Re-evaluation of the taxonomy and diversity of Russula section Foetentinae (Russulales, Basidiomycota) in Korea. Mycoscience 2017, 58, 351–360. [Google Scholar] [CrossRef]
- Adamčík, S.; Looney, B.; Caboň, M.; Jančovičová, S.; Adamčíková, K.; Avis, P.G.; Barajas, M.; Bhatt, R.P.; Corrales, A.; Das, K.; et al. The quest for a globally comprehensible Russula language. Fungal Divers. 2019, 99, 369–449. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Voucher | Location | GenBank Accession Number | |||
|---|---|---|---|---|---|---|
| ITS | nrLSU | RPB2 | TEF1 | |||
| L. albocarneus | - | China | KX441117 | KX441364 | KX442105 | - |
| L. aurantiozonatus | HCCN10589 | Korea | MH984993 | MH985118 | MH936950 | - |
| L. aurantiozonatus | SFC20150901-06 | Korea | MH984976 | MH985097 | MH936929 | - |
| L. austroscrobiculatus | MHHNU 31161 | China | OL770185 | - | - | - |
| L. austroscrobiculatus | XHW3542 | China | OL770183 | - | - | - |
| L. blennius | MTB16 | Germany | MN947353 | - | - | - |
| L. blennius | - | Sweden | AY606944 | - | - | - |
| L. brachycystidiatus | KUN-HKAS 67933 | China | MF508951 | - | - | - |
| L. brachycystidiatus | KUN-HKAS 96483 | China | MF508950 | - | - | - |
| L. brachycystidiatus | KUN-HKAS 96442 | China | MF508949 | - | - | - |
| L. brachycystidiatus | KUN-HKAS 69233 | China | MF508948 | - | - | - |
| L. brachycystidiatus | KUN-HKAS 76061 | China | MF508947 | - | - | - |
| L. brachycystidiatus | KUN-HKAS69519 | China | MF508952 | - | - | - |
| L. castaneus | KUN-HKAS 47648 | China | MF508962 | - | - | - |
| L. castaneus | KUN-HKAS 61976 | China | MF508961 | - | - | - |
| L. cinereus | LM4663F | Mexico | FJ348708 | - | - | - |
| L. cinereus | CCB9 (TENN) | USA | MF755272 | - | - | - |
| L. cinereus | 2636 | Canada | KJ705204 | - | - | - |
| L. cremicolor | KA12-0268 | Korea | MH985013 | - | QCH40198 | - |
| L. cremicolor | HCCN19933 | Korea | MH984972 | - | QCH40153 | - |
| L. dilutisalmoneus | HMJAU 42465 | China | MF152847 | - | - | - |
| L. dilutisalmoneus | KUN-HKAS 61803 | China | MF152846 | - | - | - |
| L. fluens | IBL83 | Poland | MZ410712 | - | - | - |
| L. fluens | MC01-525 | Denmark | AJ889961 | - | - | - |
| L. gloecorpus | LH91 | USA | GQ268638 | - | - | - |
| L. gloeocarpus | KUN-HKAS 73602 | China | OL770166 | - | - | - |
| L. gloeocarpus | ZP-2287 (MHHNU) | China | OL770165 | - | - | - |
| L. gloeocarpus | Pa1-mOTU056 | Japan | LC315865 | - | - | - |
| L. haugiae | LM4994 | Mexico | KT583642 | - | AOF41440 | - |
| L. haugiae | LM4988 | Mexico | KT583641 | - | AOF41439 | - |
| L. haugiae | LM4957 | Mexico | KT583640 | - | - | - |
| L. hunanensis | MHHNU 31989B | China | OL770172 | - | - | - |
| L. hunanensis | MHHNU 31989A | China | OL770171 | - | - | - |
| L. incrustatus | KUN-HKAS 61789 | China | MK675285 | - | - | - |
| L. incrustatus | KUN-HKAS 61772 | China | MK675284 | - | - | - |
| L. jianyangensis | HMAS298103T | China | OR835448 | OR826782 | OR915862 | OR887738 |
| L. jianyangensis | HMAS298104 | China | OR835446 | PP033514 | OR915863 | OR887739 |
| L. lignicola | KUN-HKAS 59474 | China | MF508946 | - | - | - |
| L. lignicola | KUN-HKAS 54122 | China | MF508945 | - | - | - |
| L. olivinus | KUN-HKAS 116928 | China | OL770196 | - | - | - |
| L. olivinus | KUN-HKAS 118988 | China | OL770195 | - | - | - |
| L. pallidizonatus | KUN-HKAS 85389 | China | MF508932 | - | - | - |
| L. pallidizonatus | KUN-HKAS 62019 | China | MF508931 | - | - | - |
| L. pallidizonatus | KUN-HKAS 85388 | China | MF508933 | - | - | - |
| L. pallidizonatus | KUN-HKAS 47641 | China | MF508930 | - | - | - |
| L. pallido-ochraceus | KUN-HKAS 71574 | China | MF508942 | - | - | - |
| L. pallido-ochraceus | KUN-HKAS 54492 | China | MF508941 | - | - | - |
| L. pallido-ochraceus | KUN-HKAS 58675 | China | MF508940 | - | - | - |
| L. pallido-ochraceus | TC_277 | China | MW722813 | - | - | - |
| L. pallido-ochraceus | KUN-HKAS 59446 | China | MF508943 | - | - | - |
| L. pallido-ochraceus | KUN-HKAS 75991 | China | MF508939 | - | - | - |
| L. parallelus | SFC20150701-55 | Korea | MH984953 | MH985072 | MH936904 | - |
| L. parallelus | KA12-1557 | Korea | MH984921 | MH985035 | MH936867 | - |
| L. pohangensis | SFC20130807-68 | Korea | MH985018 | MH985143 | MH936975 | - |
| L. pohangensis | SN1-3870 | China | LC622651 | - | - | - |
| L. psammicola | 263825 | USA | MK607513 | - | - | - |
| L. psammicola | BPL869 | USA | KY848507 | - | - | - |
| L. purpureocastaneus | KUN-HKAS 61975 | China | MF508965 | - | - | - |
| L. purpureocastaneus | KUN-HKAS 56384 | China | MF508964 | - | - | - |
| L. purpureocastaneus | KUN-HKAS 49827 | China | MF508963 | - | - | - |
| L. scrobiculatus | QHU20107 | China | OM970920 | - | - | - |
| L. scrobiculatus | HMAS264940 | China | KX441085 | KX441332 | KX442073 | - |
| L. scrobiculatus | JN01-058 | Thailand | KF432968 | - | - | - |
| L. scrobiculatus | HMAS267747 | China | KX441098 | MF893430 | ||
| L. sinozonarius | KUN-HKAS 69414 | China | MF508926 | - | - | - |
| L. sinozonarius | KUN-HKAS 56133 | China | MF508929 | - | - | - |
| L. sinozonarius | KUN-HKAS 48534 | China | MF508928 | - | 0 | - |
| L. sinozonarius | KUN-HKAS 96778 | China | MF508927 | - | - | - |
| L. sinozonarius | KUN-HKAS 54620 | China | MF508925 | - | - | - |
| L. spadiceus | SFC20121005-03 | Korea | MH985021 | MH985146 | MH936978 | - |
| L. spadiceus | NIBRFG0000104942 | Korea | MH984956 | MH985076 | MH936908 | - |
| L. subbrevipes | KUN-HKAS 85379 | China | MF508938 | - | - | - |
| L. subbrevipes | KUN-HKAS 61409 | China | MF508937 | - | - | - |
| L. subbrevipes | KUN-HKAS 57562 | China | MF508936 | - | - | - |
| L. subbrevipes | KUN-HKAS 62032 | China | MF508935 | - | - | - |
| L. subbrevipes | KUN-HKAS 94947 | China | MF508934 | - | - | - |
| L. subzonarius | KUN-HKAS 96630 | China | MF508960 | - | - | - |
| L. subzonarius | KUN-HKAS 73522 | China | MF508959 | - | - | - |
| L. uvidus | mh0963 | Sweden | AY606957 | AF325293 | - | - |
| L. uvidus | HMAS269338 | China | KX441140 | KX441387 | KX442128 | - |
| L. vulgaris | MHHNU 32017 | China | OL770178 | - | - | - |
| L. vulgaris | HYJ1878 (EFHAAU 476) | China | OL770175 | - | - | - |
| L. vulgaris | WNW33 (EFHAAU 1602) | China | OL770173 | - | - | - |
| Multifurca zonaria | FH12-009 | Thailand | KF432960 | - | - | - |
| Taxon | Voucher | Location | GenBank Accession Number | ||||
|---|---|---|---|---|---|---|---|
| ITS | nrLSU | RPB2 | mtSSU | TEF1 | |||
| R. adusta | RDL 18-039 | Belgium | OM833079 | - | - | - | ON015965.1 |
| R. aeruginea | AT2003017 | Sweden | DQ421999 | DQ421999 | - | - | - |
| R. alboareolata | SUT-1 | Thailand | AF345247 | - | - | - | - |
| R. albolutea | RITF4460 | China: Chongqing | - | MW397121 | MW411341 | MW403834 | - |
| R. albolutea | RITF4461 | China: Yunnan | - | MW397122 | MW411342 | MW403835 | - |
| R. albolutea | RITF4462 | China: Yunnan | - | MW397123 | MW411343 | MW403836 | - |
| R. albolutea | RITF2653 | China | MT672478 | MW397120 | MW411340 | MW403833 | - |
| R. amethystina | SJ84 | Pakistan | KT953615 | - | - | - | - |
| R. amethystina | SJ92 | Pakistan | KT953616 | - | - | - | - |
| R. amoenolens | 12838 (MICH) | France | KF245510 | - | - | - | - |
| R. amoenolens | 77763 (PDD) | New Zealand | GU222264 | - | - | - | - |
| R. ammophila | IC09010703 (BCN) | Spain | MK112566 | MK108033 | - | - | - |
| R. aureoviridis | H15060613 | China | KY767808 | - | - | - | - |
| R. aureoviridis | RITF1868 | China | MW397096 | - | - | MW403842 | - |
| R. aureoviridis | H16082612 | China | KY767809 | MK881920 | - | MK882048 | MN617846 |
| R. aureoviridis | RITF4709 | China | MW646980 | MW646992 | - | MW647003 | MW650849 |
| R. brunneoaurantiaca | GUBH19957 | India | OP270714 | - | - | - | - |
| R. brunneonigra | H5813 (DAR) | Australia | EU019945 | - | - | - | - |
| R. cerolens | HE2720 | China | KC505578 | - | - | - | - |
| R. cf. amoenolens | 12794 (MICH) | USA | KF245512 | - | - | - | - |
| R. cf. vesca | BB 06.525 | Mexico | - | KU237465 | KU237751 | KU237309 | - |
| R. crassotunicata | UBCOGTR0439s | Canada | EU597082 | - | - | - | - |
| R. crustosa | BPL265 | USA: Tennessee | - | KT933826 | KT933898 | - | - |
| R. cyanoxantha | UE29.09.2002-2 | France | - | DQ422033 | DQ421970 | - | - |
| R. cyanoxantha | FH 12-201 | Germany | KR364093 | KR364225 | - | - | - |
| R. farinipes | UE28.09.2002-4 | Sweden | DQ421983 | - | - | - | - |
| R. fuliginosa | R74 | CZECH REPUBLIC | HG798529 | - | - | - | - |
| R. grisea | BB 07.184 | Slovakia | - | KU237509 | KU237795 | KU237355 | - |
| R. grisea | UE2005.08.16-01 | Sweden | DQ422030 | DQ422030 | - | - | - |
| R. grisea | FH12234 | Germany | KT934006 | KT933867 | - | - | - |
| R. ilicis | MF 00.300 | Italy | - | KU237595 | KU237880 | KU237443 | - |
| R. illota | UE26.07.2002-3 (UPS) | Sweden | DQ422024 | DQ422024 | DQ421967 | - | - |
| R. indoalba | AG 15-628 | India | KX234820 | - | - | - | - |
| R. junzifengensis | HMAS298101T | China | OR826832 | OR826833 | OR915864 | OR941507 | OR887742 |
| R. junzifengensis | HMAS298102 | China | OR880061 | OR880054 | OR915865 | OR941508 | OR887743 |
| R. langei | BB 07.792 | France | - | KU237510 | KU237796 | KU237356 | - |
| R. livescens | F0177 | China | GU371295 | - | - | - | - |
| R. maguanensis | XHW4765 | China | MH724918 | MH714537 | MH939990 | - | MH939983 |
| R. mariae | BB 07.038 | USA | - | KU237538 | KU237824 | KU237384 | - |
| R. medullata | BB 07.252 | Slovakia | - | KU237546 | KU237832 | KU237392 | - |
| R. mustelina | FH12226 | Germany | - | KT933866 | KT933937 | - | - |
| R. mustelina | SA 09.88 | Slovakia | - | KU237596 | KU237881 | KU237444 | - |
| R. nigricans | - | Germany | AF418607 | - | - | - | - |
| R. pallescens | UBC:F23913 | Canada | KJ146729 | - | - | - | - |
| R. pallidula | RITF2613 | China | - | MH027960 | MH091698 | MW403845 | - |
| R. pallidula | RITF3331 | China | - | MH027961 | MH091699 | MW403846 | - |
| R. parvovirescens | SDRM 6280 | USA | MK532789 | - | - | - | - |
| R. prasina | HMAS 281232 | China | MH454351 | - | - | - | - |
| R. pseudocrustosa | HBAU15015 | China | MT337520 | - | - | - | - |
| R. pulverulenta | RF566 (pers. herb.) | USA | AY061736 | - | - | - | - |
| R. rufobasalis | H17052204 | China | MH168570 | MK881947.1 | - | MK882075.1 | MT085585.1 |
| R. subpunicea | RITF1435 | China: Hunan | - | MW397126 | MW411346 | MW403839 | - |
| R. subpunicea | RITF2615 | China: Hunan | - | MW397127 | MW411347 | MW403840 | - |
| R. subpunicea | RITF3715 | China | MN833635 | MW397124 | MW411344 | MW403837 | - |
| R. subpunicea | RITF2648 | China | MN833638 | MW397125 | MW411345 | MW403838 | - |
| R. substriata | XHW4766 | China | MH724921 | MH714540 | MH939993 | - | MH939986 |
| R. variata | BPL241 | USA | - | KT933818 | KT933889 | - | - |
| R. vesca | BPL284 | USA | KT933978 | KT933839 | - | - | - |
| R. vesca | AT2002091 | Sweden | DQ422018 | DQ422018 | DQ421959 | - | - |
| R. violeipes | BB 07.273 | Slovakia | KU237534 | KU237820 | KU237380 | - | |
| R. virescens | HJB9989 | Belgium | DQ422014 | DQ421955 | - | - | |
| R. xanthovirens | GDGM 71145 | China | MG786056 | - | - | - | - |
| R. zonatus | HMAS298099 | China | OR826839 | OR826846 | OR915866 | OR941505 | OR887740 |
| R. zonatus | HMAS298100T | China | OR880062 | OR880056 | OR915867 | OR941506 | OR887741 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Zhu, M.; Keyhani, N.O.; Wu, Z.; Lv, H.; Heng, Z.; Chen, R.; Dang, Y.; Yang, C.; Chen, J.; et al. Three New Species of Russulaceae (Russulales, Basidiomycota) from Southern China. J. Fungi 2024, 10, 70. https://doi.org/10.3390/jof10010070
Liu S, Zhu M, Keyhani NO, Wu Z, Lv H, Heng Z, Chen R, Dang Y, Yang C, Chen J, et al. Three New Species of Russulaceae (Russulales, Basidiomycota) from Southern China. Journal of Fungi. 2024; 10(1):70. https://doi.org/10.3390/jof10010070
Chicago/Turabian StyleLiu, Sen, Mengjia Zhu, Nemat O. Keyhani, Ziyi Wu, Huajun Lv, Zhiang Heng, Ruiya Chen, Yuxiao Dang, Chenjie Yang, Jinhui Chen, and et al. 2024. "Three New Species of Russulaceae (Russulales, Basidiomycota) from Southern China" Journal of Fungi 10, no. 1: 70. https://doi.org/10.3390/jof10010070