
Ritonavir Has Reproductive Toxicity Depending on Disrupting PI3K/PDK1/AKT Signaling Pathway

Abstract
:1. Introduction
2. Materials and Methods
2.1. Media and Chemicals
2.2. Sample Preparation
2.3. Sperm Motility and Motion Kinematics
2.4. Sperm Capacitation Status
2.5. Cell Viability
2.6. Western Blotting Analysis
2.7. Statistical Analysis
3. Results
3.1. The Impact of Ritonavir on Sperm Motility and Motion Kinematics
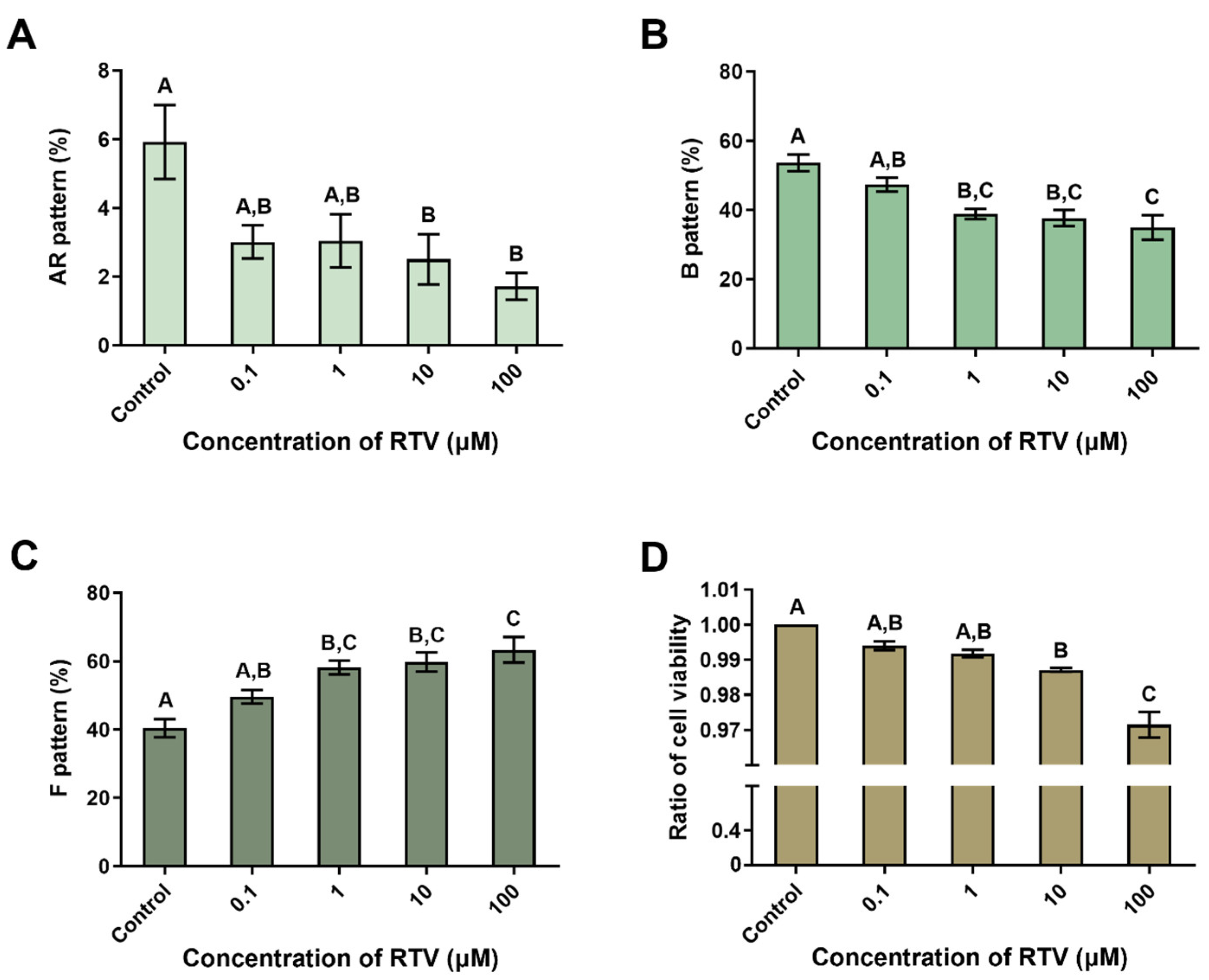
3.2. The Impact of Ritonavir on Capacitation Status and Cell Viability
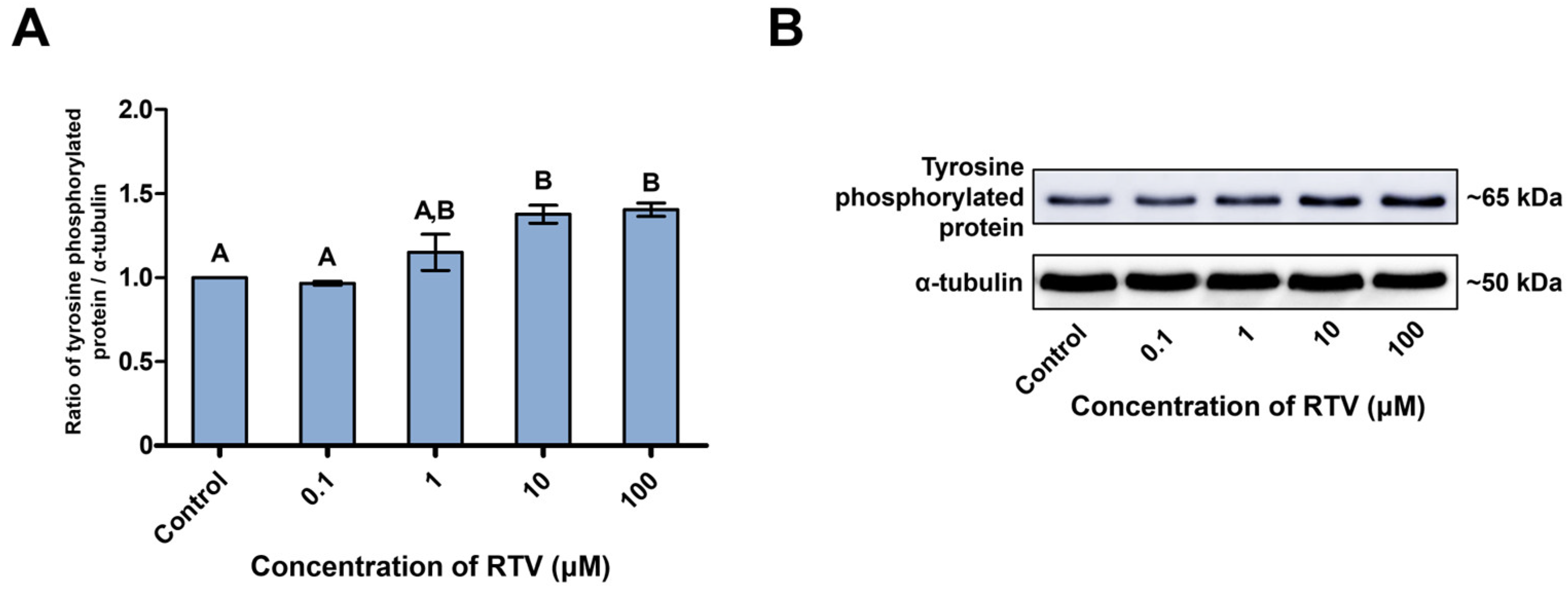
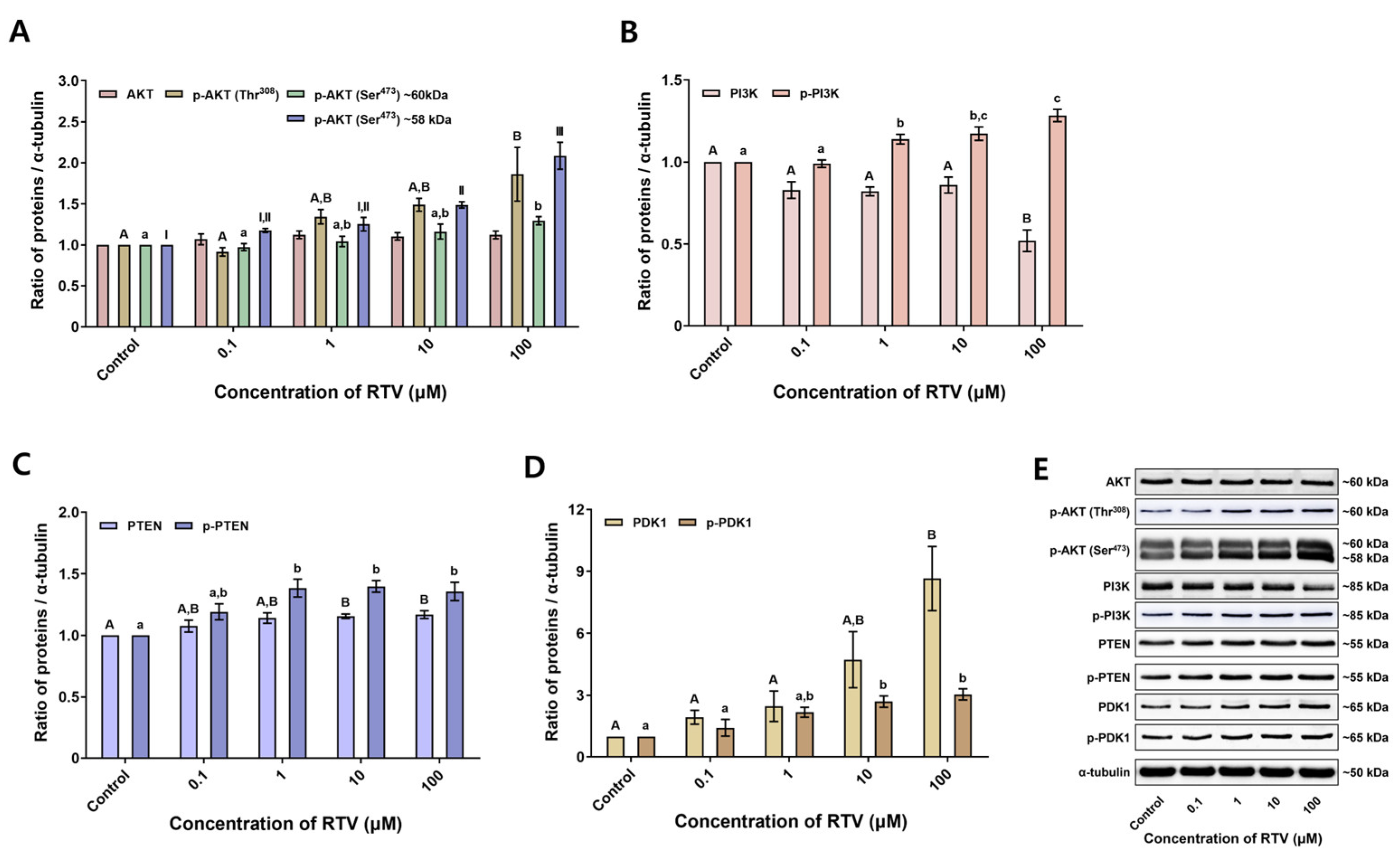
3.3. Expression Levels of Tyrosine-Phosphorylated Proteins and PI3K/PDK1/AKT Pathway-Related Proteins
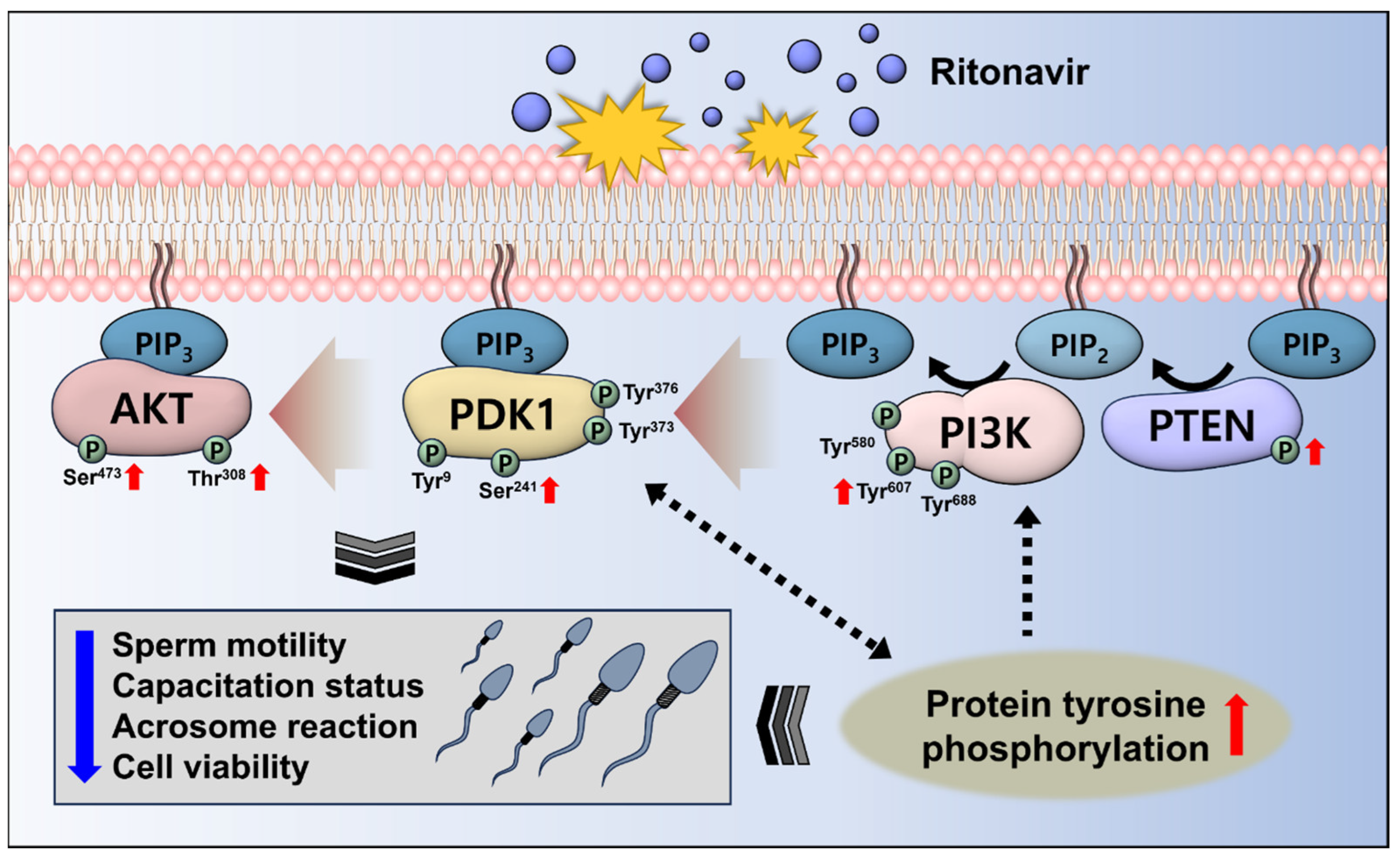
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kempf, D.J.; Marsh, K.C.; Denissen, J.F.; McDonald, E.; Vasavanonda, S.; Flentge, C.A.; Green, B.E.; Fino, L.; Park, C.H.; Kong, X.-P. ABT-538 is a potent inhibitor of human immunodeficiency virus protease and has high oral bioavailability in humans. Proc. Natl. Acad. Sci. USA 1995, 92, 2484–2488. [Google Scholar] [CrossRef]
- Martinez, M.A. Compounds with therapeutic potential against novel respiratory 2019 coronavirus. Antimicrob. Agents Chemother. 2020, 64, e00399-20. [Google Scholar] [CrossRef] [PubMed]
- Baden, L.R.; Rubin, E.J. COVID-19—The search for effective therapy. N. Engl. J. Med. 2020, 382, 1851–1852. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Cao, R.; Zhang, L.; Yang, X.; Liu, J.; Xu, M.; Shi, Z.; Hu, Z.; Zhong, W.; Xiao, G. Remdesivir and chloroquine effectively inhibit the recently emerged novel coronavirus (2019-nCoV) in vitro. Cell Res. 2020, 30, 269–271. [Google Scholar] [CrossRef]
- Mahase, E. COVID-19: Pfizer’s paxlovid is 89% effective in patients at risk of serious illness, company reports. BMJ 2021, 375, n2713. [Google Scholar] [CrossRef]
- Srirangam, A.; Mitra, R.; Wang, M.; Gorski, J.C.; Badve, S.; Baldridge, L.; Hamilton, J.; Kishimoto, H.; Hawes, J.; Li, L.; et al. Effects of HIV protease inhibitor ritonavir on Akt-regulated cell proliferation in breast cancer. Clin. Cancer Res. 2006, 12, 1883–1896. [Google Scholar] [CrossRef]
- Batchu, R.B.; Gruzdyn, O.V.; Bryant, C.S.; Qazi, A.M.; Kumar, S.; Chamala, S.; Kung, S.T.; Sanka, R.S.; Puttagunta, U.S.; Weaver, D.W.; et al. Ritonavir-Mediated Induction of Apoptosis in Pancreatic Cancer Occurs via the RB/E2F-1 and AKT Pathways. Pharmaceuticals 2014, 7, 46–57. [Google Scholar] [CrossRef]
- Zhong, D.S.; Lu, X.H.; Conklin, B.S.; Lin, P.H.; Lumsden, A.B.; Yao, Q.; Chen, C. HIV protease inhibitor ritonavir induces cytotoxicity of human endothelial cells. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 1560–1566. [Google Scholar] [CrossRef]
- Ganta, K.K.; Chaubey, B. Endoplasmic reticulum stress leads to mitochondria-mediated apoptosis in cells treated with anti-HIV protease inhibitor ritonavir. Cell Biol. Toxicol. 2019, 35, 189–204. [Google Scholar] [CrossRef]
- Jung, E.-J.; Lee, W.-J.; Hwang, J.-M.; Bae, J.-W.; Kwon, W.-S. Reproductive Toxicity of Ritonavir in Male: Insight into mouse sperm capacitation. Reprod. Toxicol. 2022, 114, 1–6. [Google Scholar] [CrossRef]
- Austin, C.R. The ‘capacitation’ of the mammalian sperm. Nature 1952, 170, 326. [Google Scholar] [CrossRef]
- Austin, C. Observations on the penetration of the sperm into the mammalian egg. Aust. J. Biol. Sci. 1951, 4, 581–596. [Google Scholar] [CrossRef]
- Chang, M.C. Fertilizing capacity of spermatozoa deposited into the fallopian tubes. Nature 1951, 168, 697–698. [Google Scholar] [CrossRef]
- Breitbart, H.; Rotman, T.; Rubinstein, S.; Etkovitz, N. Role and regulation of PI3K in sperm capacitation and the acrosome reaction. Mol. Cell. Endocrinol. 2010, 314, 234–238. [Google Scholar] [CrossRef]
- Fisher, H.; Brewis, I.A.; Barratt, C.; Cooke, I.; Moore, H. Phosphoinositide 3-kinase is involved in the induction of the human sperm acrosome reaction downstream of tyrosine phosphorylation. Mol. Hum. Reprod. 1998, 4, 849–855. [Google Scholar] [CrossRef]
- Koppers, A.J.; Mitchell, L.A.; Wang, P.; Lin, M.; Aitken, R.J. Phosphoinositide 3-kinase signalling pathway involvement in a truncated apoptotic cascade associated with motility loss and oxidative DNA damage in human spermatozoa. Biochem. J. 2011, 436, 687–698. [Google Scholar] [CrossRef]
- Jang, S.-I.; Jo, J.-H.; Jung, E.-J.; Lee, W.-J.; Hwang, J.-M.; Bae, J.-W.; Kwon, W.-S. Development of an optimal protocol to induce capacitation of boar spermatozoa in vitro. J. Anim. Reprod. Biotechnol. 2022, 37, 285–291. [Google Scholar] [CrossRef]
- Lee, W.-J.; Jung, E.-J.; Hwang, J.-M.; Bae, J.-W.; Kwon, W.-S. GRP78 plays a key role in sperm function via the PI3K/PDK1/AKT pathway. Reprod. Toxicol. 2022, 113, 103–109. [Google Scholar] [CrossRef]
- Lee, W.-J.; Jo, J.-H.; Jang, S.-I.; Jung, E.-J.; Hwang, J.-M.; Bae, J.-W.; Ha, J.J.; Kim, D.-H.; Kwon, W.-S. The Natural Flavonoid Compound Deguelin Suppresses Sperm (Sus Scrofa) Functions through Abnormal Activation of the PI3K/AKT Pathway. Reprod. Toxicol. 2023, 120, 108426. [Google Scholar] [CrossRef]
- Arend, C.; Brandmann, M.; Dringen, R. The antiretroviral protease inhibitor ritonavir accelerates glutathione export from cultured primary astrocytes. Neurochem. Res. 2013, 38, 732–741. [Google Scholar] [CrossRef] [PubMed]
- Lan, X.; Kiyota, T.; Hanamsagar, R.; Huang, Y.; Andrews, S.; Peng, H.; Zheng, J.C.; Swindells, S.; Carlson, G.A.; Ikezu, T. The effect of HIV protease inhibitors on amyloid-β peptide degradation and synthesis in human cells and Alzheimer’s disease animal model. J. Neuroimmune Pharmacol. 2012, 7, 412–423. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.-M.; Bae, J.-W.; Jung, E.-J.; Lee, W.-J.; Kwon, W.-S. Novaluron Has Detrimental Effects on Sperm Functions. Int. J. Environ. Res. Public Health 2021, 19, 61. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-H.; Bae, J.-W.; Kim, D.-H.; Jeong, D.-J.; Ha, J.J.; Yi, J.K.; Kwon, W.-S. Detrimental effects of temephos on male fertility: An in vitro study on a mouse model. Reprod. Toxicol. 2020, 96, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.-W.; Kwon, W.-S. Investigating the effects of fipronil on male fertility: Insight into the mechanism of capacitation. Reprod. Toxicol. 2020, 94, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Marzolini, C.; Kuritzkes, D.R.; Marra, F.; Boyle, A.; Gibbons, S.; Flexner, C.; Pozniak, A.; Boffito, M.; Waters, L.; Burger, D. Recommendations for the management of drug–drug interactions between the COVID-19 antiviral nirmatrelvir/ritonavir (Paxlovid) and comedications. Clin. Pharmacol. Ther. 2022, 112, 1191–1200. [Google Scholar] [CrossRef]
- Wang, M.W.-H.; Wei, S.; Faccio, R.; Takeshita, S.; Tebas, P.; Powderly, W.G.; Teitelbaum, S.L.; Ross, F.P. The HIV protease inhibitor ritonavir blocks osteoclastogenesis and function by impairing RANKL-induced signaling. J. Clin. Investig. 2004, 114, 206–213. [Google Scholar] [CrossRef]
- Dalva-Aydemir, S.; Bajpai, R.; Martinez, M.; Adekola, K.U.; Kandela, I.; Wei, C.; Singhal, S.; Koblinski, J.E.; Raje, N.S.; Rosen, S.T.; et al. Targeting the metabolic plasticity of multiple myeloma with FDA-approved ritonavir and metformin. Clin. Cancer Res. 2015, 21, 1161–1171. [Google Scholar] [CrossRef]
- Kumar, S.; Bryant, C.S.; Chamala, S.; Qazi, A.; Seward, S.; Pal, J.; Steffes, C.P.; Weaver, D.W.; Morris, R.; Malone, J.M.; et al. Ritonavir blocks AKT signaling, activates apoptosis and inhibits migration and invasion in ovarian cancer cells. Mol. Cancer 2009, 8, 26. [Google Scholar] [CrossRef]
- Stambolic, V.; Suzuki, A.; De La Pompa, J.L.; Brothers, G.M.; Mirtsos, C.; Sasaki, T.; Ruland, J.; Penninger, J.M.; Siderovski, D.P.; Mak, T.W. Negative regulation of PKB/Akt-dependent cell survival by the tumor suppressor PTEN. Cell 1998, 95, 29–39. [Google Scholar] [CrossRef]
- Alessi, D.R.; James, S.R.; Downes, C.P.; Holmes, A.B.; Gaffney, P.R.; Reese, C.B.; Cohen, P. Characterization of a 3-phosphoinositide-dependent protein kinase which phosphorylates and activates protein kinase Bα. Curr. Biol. 1997, 7, 261–269. [Google Scholar] [CrossRef]
- Alessi, D.R.; Andjelkovic, M.; Caudwell, B.; Cron, P.; Morrice, N.; Cohen, P.; Hemmings, B.A. Mechanism of activation of protein kinase B by insulin and IGF-1. EMBO J. 1996, 15, 6541–6551. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, W.-D.; Liu, X.-Q.; Zhang, P.-F.; Hao, Y.-N.; Li, L.; Chen, L.; Shen, W.; Tang, X.-F.; Min, L.-J. Hydrogen sulfide and/or ammonia reduces spermatozoa motility through AMPK/AKT related pathways. Sci. Rep. 2016, 6, 37884. [Google Scholar] [CrossRef]
- Dorostghoal, M.; Galehdari, H.; Moramezi, F.; Danyari, R. Sperm miR-26a-5p and its target PTEN transcripts content in men with unexplained infertility. Andrology 2020, 8, 1167–1173. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Fan, Y.; Zhang, J.; Feng, S.; Hu, Z.; Qiu, W.; Long, K.; Jin, L.; Tang, Q.; Wang, X. Testosterone-dependent miR-26a-5p and let-7g-5p act as signaling mediators to regulate sperm apoptosis via targeting PTEN and PMAIP1. Int. J. Mol. Sci. 2018, 19, 1233. [Google Scholar] [CrossRef] [PubMed]
- Aparicio, I.; Bragado, M.; Gil, M.; Garcia-Herreros, M.; Gonzalez-Fernandez, L.; Tapia, J.; Garcia-Marin, L. Phosphatidylinositol 3-kinase pathway regulates sperm viability but not capacitation on boar spermatozoa. Mol. Reprod. Dev. 2007, 74, 1035–1042. [Google Scholar] [CrossRef] [PubMed]
- Li, X.H.; Chai, R.R.; Chen, G.W.; Zhang, L.F.; Tan-Tai, W.J.; Shi, H.J.; Martin-DeLeon, P.A.; O, W.S.; Chen, H. Prohibitin (PHB) interacts with AKT in mitochondria to coordinately modulate sperm motility. Asian J. Androl. 2020, 22, 583–589. [Google Scholar] [CrossRef]
- Zhang, T.-Y.; Wu, R.-Y.; Zhao, Y.; Xu, C.-S.; Zhang, W.-D.; Ge, W.; Liu, J.; Sun, Z.-Y.; Zou, S.-H.; Shen, W. Ochratoxin A exposure decreased sperm motility via the AMPK and PTEN signaling pathways. Toxicol. Appl. Pharmacol. 2018, 340, 49–57. [Google Scholar] [CrossRef]
- Gallardo Bolanos, J.M.; Balao da Silva, C.M.; Martin Munoz, P.; Morillo Rodriguez, A.; Plaza Davila, M.; Rodriguez-Martinez, H.; Aparicio, I.M.; Tapia, J.A.; Ortega Ferrusola, C.; Pena, F.J. Phosphorylated AKT preserves stallion sperm viability and motility by inhibiting caspases 3 and 7. Reproduction 2014, 148, 221–235. [Google Scholar] [CrossRef]
- Quan, Y.; Liu, Q. Effect of Akti-2 on sperm motility, capacitation and acrosome reaction in a mouse model. Biomed. Rep. 2016, 4, 578–582. [Google Scholar] [CrossRef]
- Macías-García, B.; García-Marín, L.J.; Bragado, M.J.; González-Fernández, L. The calcium-sensing receptor regulates protein tyrosine phosphorylation through PDK1 in boar spermatozoa. Mol. Reprod. Dev. 2019, 86, 751–761. [Google Scholar] [CrossRef]
- Harayama, H.; Nakamura, K. Changes of PKA and PDK1 in the principal piece of boar spermatozoa treated with a cell-permeable cAMP analog to induce flagellar hyperactivation. Mol. Reprod. Dev. 2008, 75, 1396–1407. [Google Scholar] [CrossRef] [PubMed]
- von Willebrand, M.; Williams, S.; Saxena, M.; Gilman, J.; Tailor, P.; Jascur, T.; Amarante-Mendes, G.P.; Green, D.R.; Mustelin, T. Modification of phosphatidylinositol 3-kinase SH2 domain binding properties by Abl-or Lck-mediated tyrosine phosphorylation at Tyr-688. J. Biol. Chem. 1998, 273, 3994–4000. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Hill, M.M.; Hess, D.; Brazil, D.P.; Hofsteenge, J.; Hemmings, B.A. Identification of tyrosine phosphorylation sites on 3-phosphoinositide-dependent protein kinase-1 and their role in regulating kinase activity. J. Biol. Chem. 2001, 276, 37459–37471. [Google Scholar] [CrossRef] [PubMed]
- Grillo, S.; Grémeaux, T.; Casamayor, A.; Alessi, D.R.; Le Marchand-Brustel, Y.; Tanti, J.F. Peroxovanadate induces tyrosine phosphorylation of phosphoinositide-dependent protein kinase-1: Potential involvement of Src kinase. Eur. J. Biochem. 2000, 267, 6642–6649. [Google Scholar] [CrossRef]
- Yang, K.-J.; Shin, S.; Piao, L.; Shin, E.; Li, Y.; Park, K.A.; Byun, H.S.; Won, M.; Hong, J.; Kweon, G.R. Regulation of 3-phosphoinositide-dependent protein kinase-1 (PDK1) by Src involves tyrosine phosphorylation of PDK1 and Src homology 2 domain binding. J. Biol. Chem. 2008, 283, 1480–1491. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration of RTV (µM) | |||||
|---|---|---|---|---|---|
| Control | 0.1 | 1 | 10 | 100 | |
| MOT (%) | 77.87 ± 1.60 A | 78.48 ± 2.22 A | 76.73 ± 2.13 A | 71.86 ± 2.61 A | 45.63 ± 2.97 B |
| RPD (%) | 65.47 ± 2.36 A | 65.19 ± 2.92 A | 62.78 ± 2.69 A | 56.60 ± 3.77 A | 32.20 ± 2.38 B |
| MED (%) | 8.19 ± 0.67 | 8.83 ± 0.70 | 8.37 ± 0.71 | 9.48 ± 0.73 | 8.62 ± 0.57 |
| SLW (%) | 3.90 ± 0.58 | 4.08 ± 0.53 | 5.25 ± 0.80 | 5.66 ± 0.95 | 4.47 ± 0.47 |
| PRG (%) | 73.66 ± 2.07 A | 74.02 ± 2.64 A | 71.15 ± 2.49 A | 66.08 ± 3.24 A | 40.82 ± 2.91 B |
| VCL (μm/s) | 84.87 ± 3.66 A | 87.52 ± 5.18 A | 80.91 ± 3.90 A | 71.45 ± 4.78 A | 38.49 ± 3.23 B |
| VSL (μm/s) | 45.95 ± 2.09 A | 44.60 ± 2.01 A | 42.00 ± 2.14 A | 39.63 ± 2.25 A | 20.87 ± 1.31 B |
| VAP (μm/s) | 60.58 ± 2.81 A | 60.14 ± 3.29 A | 55.57 ± 3.00 A | 50.68 ± 3.52 A | 26.76 ± 2.09 B |
| LIN (%) | 42.96 ± 1.67 A | 40.73 ± 1.35 A | 40.10 ± 1.73 A | 39.74 ± 1.35 A | 24.96 ± 1.30 B |
| STR (%) | 76.22 ± 2.18 | 74.54 ± 1.69 | 76.00 ± 1.85 | 78.79 ± 1.59 | 78.72 ± 1.80 |
| BCF (Hz) | 6.59 ± 0.26 A | 6.54 ± 0.30 A | 6.48 ± 0.22 A | 5.83 ± 0.33 A | 2.78 ± 0.18 B |
| MAD (degree) | 64.57 ± 2.91 A | 63.42 ± 2.37 A | 59.78 ± 2.50 A | 57.31 ± 2.77 A | 31.49 ± 1.69 B |
| WOB (%) | 71.49 ± 1.43 | 69.12 ± 1.30 | 68.66 ± 1.22 | 70.87 ± 0.62 | 69.84 ± 1.10 |
| DNC (μm2/s) | 261.99 ± 24.25 A | 291.66 ± 37.26 A | 248.29 ± 24.44 A | 190.24 ± 26.57 A | 62.92 ± 11.47 B |
| DNM (μm) | 7.12 ± 0.51 A,B | 7.96 ± 0.58 A | 7.50 ± 0.50 A,B | 6.29 ± 0.33 A,B | 5.78 ± 0.40 B |
| ALH (μm) | 2.98 ± 0.17 A | 3.17 ± 0.22 A | 2.94 ± 0.16 A | 2.49 ± 0.19 A | 1.43 ± 0.15 B |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, E.-J.; Jo, J.-H.; Uwamahoro, C.; Jang, S.-I.; Lee, W.-J.; Hwang, J.-M.; Bae, J.-W.; Kwon, W.-S. Ritonavir Has Reproductive Toxicity Depending on Disrupting PI3K/PDK1/AKT Signaling Pathway. Toxics 2024, 12, 73. https://doi.org/10.3390/toxics12010073
Jung E-J, Jo J-H, Uwamahoro C, Jang S-I, Lee W-J, Hwang J-M, Bae J-W, Kwon W-S. Ritonavir Has Reproductive Toxicity Depending on Disrupting PI3K/PDK1/AKT Signaling Pathway. Toxics. 2024; 12(1):73. https://doi.org/10.3390/toxics12010073
Chicago/Turabian StyleJung, Eun-Ju, Jae-Hwan Jo, Claudine Uwamahoro, Seung-Ik Jang, Woo-Jin Lee, Ju-Mi Hwang, Jeong-Won Bae, and Woo-Sung Kwon. 2024. "Ritonavir Has Reproductive Toxicity Depending on Disrupting PI3K/PDK1/AKT Signaling Pathway" Toxics 12, no. 1: 73. https://doi.org/10.3390/toxics12010073